Chapter: Genetics and Molecular Biology: Genetics
Classical Genetics of Chromosomes
Classical Genetics of Chromosomes
We should not proceed to a detailed discussion of
molecular genetics without a brief review of Mendelian genetics. Chromosomes in
eukary-otes consist mainly of DNA and histones. During some stages of the
cell’s division cycle in plants and animals, chromosomes can be observed with
light microscopes, and they display beautiful and fascinating patterns. Careful
microscopic study of such chromosomes sets the stage for subsequent molecular
experiments that have revealed the exact chemi-cal nature of heredity. We are
now approaching a similar level of understanding of genetic recombination.
The basis of many of the classical studies is that
most types of eukaryotic cells are diploid. This means that each cell contains
pairs of identical or almost identical homologous chromosomes, one chromo-some
of each pair deriving from each of the parents. There are excep-tions. Some
types of plants are tetraploid or even octaploid, and some variants of other
species possess alternate numbers of one or more of the chromosomes.
During normal cell growth and division, the pairs
of chromosomes in each dividing cell are duplicated and distributed to the two
daughter cells in a process called mitosis. As a result, each daughter cell
receives the same genetic information as the parent cell contained. The
situation, however, must be altered for sexual reproduction. During this
process, special cells derived from each of the parents fuse and give rise to
the new progeny. To maintain a constant amount of DNA per cell from one
generation to the next, the special cells, which are often called gametes, are
generated. These contain only one copy of each chromosome instead of the two
copies contained by most cells. The normal chromosome number of two is called
diploid, and a chromosome number of half this is called haploid. The cell
divisions giving rise to the haploid gametes in animals and haploid spores in
plants is called meiosis.
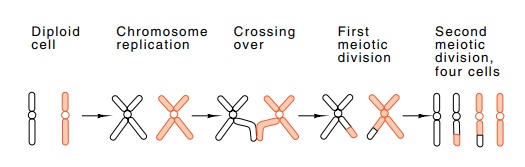
Figure
8.3 The classical view of meiosis.
During the process of meiosis, a pair of
chromosomes doubles, genetic recombination may occur between homologous
chromosomes, and the cell then divides (Fig. 8.3). Each of the daughter cells
then divides again without duplication of the chromosomes. The net result is
four cells, each containing only one copy of each chromosome. Subsequent fusion
of a sperm and egg cell from different individuals yields a diploid called a
zygote that grows and divides to yield an organism containing one member of
each chromosome pair from each parent.
The chromosomes from each of the parents may
contain mutations that produce recognizable traits or phenotypes in the
offspring. Let us consider just one chromosome pair of a hypothetical organism.
Let gene A produce trait A, and, if
it is mutant, let it be denoted as gene a
and itstrait be a. In genetic terminology, A and a are alleles. We
can describe the genetic state of an individual by giving its genetic composition
or genotype. For example, both copies of the chromosome in question could
contain the A allele. For
convenience, denote this as (A/A). Such a cell is called homozygous for
gene A. A mating between organisms
containing diploid cells of type (A/A) and (a/a) must produce
offspring of the type (a/A), which is, of course, identical to (A/a).
That is, the chromosomes in the offspring are copies of each of the parental
chro - mosomes. These offspring are said to be heterozygous for gene A.
Figure
8.4 The haploids produced from
diploid parent cells, the combinationsof haploids possible upon their fusion,
and the apparent phenotypes if A is
totally or partially dominant to a.

The interesting results come when two heterozygous
individuals mate and produce offspring. A gamete can inherit one or the other
of each of the homologous chromosomes from each chromosome pair. This generates
a variety of gamete types. When large numbers of offspring are considered, many
representatives are found of every possible combination of assortment of the
chromosomes, and the re-sults become predictable. It is easiest to systematize
the possibilities in a square matrix (Fig. 8.4). For evaluation of experimental
results, however, the appearance of heterozygotes must be known or deduced. The
appearance of a heterozygote (a/A) is that of trait A if A is dominant, which means automatically
that a is recessive. Strict dominance
need not be seen, and a heterozygote may combine the traits displayed by the
two alleles. For example, if the trait of gene A were the production of red pigment in flowers and the trait of
gene a were the absence of production
of the pigment, the heterozygote (a/A) might produce half the normal amount
of red pigment and yield pink flowers.
Related Topics