Principles of Inheritance and Variation - Extra chromosomal / cytoplasmic inheritance | 12th Zoology : Chapter 4 : Principles of Inheritance and Variation
Chapter: 12th Zoology : Chapter 4 : Principles of Inheritance and Variation
Extra chromosomal / cytoplasmic inheritance
Extra
chromosomal / cytoplasmic inheritance
Certain characters are
controlled by non-nuclear genomes found in chloroplast, mitochondria, infective
agents and plasmids. These characters do not reveal Mendelian pattern of
inheritance. The inheritance of the extra chromosomal genes are found to
exhibit maternal influence. Maternal effect is due to the asymmetric
contribution of the female parent to the development of zygote. Although both
male and female parents contribute equally to the zygote in terms of
chromosomal genes, the female parent usually contributes the zygote’s initial
cytoplasm and organelles, since the sperms contain very little cytoplasm. If
there are hereditary units in the cytoplasm, these will be transmitted to the
offsprings through the egg, so the offsprings exhibit maternal effect.
The cytoplasmic
extranuclear genes have a characteristic pattern of inheritance which do not
resemble the genes of nuclear chromosomes and is known as extra chromosomal or
extra nuclear or cytoplasmic inheritance and exhibit maternal influence. In
extra nuclear inheritance, male and female parents contribute equally their
nuclear genes to the progeny but do not make equal contribution of extra
chromosomal genes hence, the crosses can yield different (or) non Mendelian
results. Cytoplasmic inheritance in animals can be studied with reference to
shell coiling in Limnaea and kappa particles in Paramecium.
1. Shell coiling in Limnaea
Limnaea peregra is a freshwater snail.
The shell of these animals are spirally coiled. The coiling of the shell is
clockwise (dextral) or anticlockwise (sinistral). Both type of coilings are
produced by two different types of genetically controlled cleavages namely,
dextral cleavage and sinistral cleavage.
In Limnaea,
dextral coiling is normal and sinistral coiling is a mutant character.
Direction of coiling is determined by a pair of nuclear genes, D(dextral) and d
(sinistral). The gene for dextral (D) being dominant over sinistral coiling
(d). In Fig.4.11 a dextral snail provides the eggs and a sinistral snail
provides the sperm. The offsprings are all dextral (Dd), in the F1
generation.
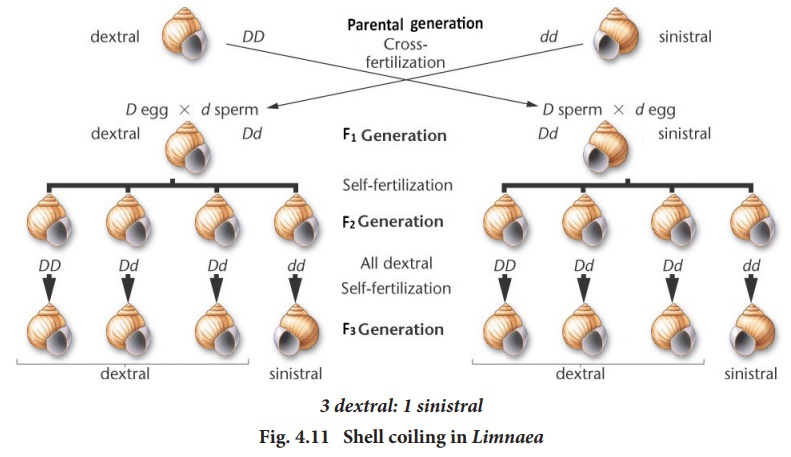
When the F1
heterozygous dextral individual (Dd) were self crossed the F2
generation showed dextral coiling with genotype of 1DD, 2Dd and 1dd (Fig-4.11
left).
When a reciprocal cross is made (Fig. 4.11-right) The F1 individuals have Dd genotype but are coiled sinistrally, as in the female parent. In both the crosses the F1 are phenotypically similar to the female parent, though the offsprings in both crosses have the same genotype Dd. This is because the genotype of the maternal parent determines the phenotype of the offspring.
When the F1
sinistral individuals were self crossed, the shell coiling in the F2
generation, were all dextral (Fig-4.11 right). This is because the genes
do not segregate in the F2 generation. Only in the F 3
generation segregation occurs in the ratio of 3 dextral : 1 sinistral.
Why does this pattern
occur? The type of cleavage depends on the organization of the egg which is
established before the maturation division of the oocyte nucleus and by the
influence of the maternal genotype. The direction of coiling of the shell
depends upon the orientation of the mitotic spindle during the first cleavage.
Obviously, maternal control affects only one generation. In each generation the
coiling is dependent on the maternal genotype.
![]()
![]()
2. Kappa particles in Paramecium
Sonneborn and his associates have reported the transmission of the cytoplasmic kappa particles in Paramecium aurelia. The kappa particles are cytoplasmic symbionts occurring in some strains of the ciliated Paramecium. The strains possessing the kappa particles are known as “killer Paramecia”.
The kappa liberates a
toxin, paramecin which is lethal to other individuals called
“sensitives”.
Kappa particles appear
to be either parasites or more possible symbionts since they do not harm their
hosts. A killer Paramecium may contain hundreds of kappa particles
which have their own DNA and which in turn are dependent on a dominant gene ‘K’
for its presence in the killer Paramecia. Paramecia with nuclear
genotype “kk” are unable to produce kappa particles. The inheritance of killer
trait does not follow the Mendelian pattern of inheritanc.
When a killer Paramecium
KK conjugates with sensitive ”kk”, the exconjugants are all heterozygous for Kk
genes. The Kk genotype suggest that both exconjugants should be killers. But
this is not seen. If conjugation lasts only for a short period of time, there
is no exchange of cytoplasm between the two Paramecia resulting in both
killers (Kk) and sensitives. However prolonged conjugation permits
mixing of cytoplasm of both the conjugants resulting in killers only. This
confirms that the killer trait is determind cytoplasmically. Dominant
chromosomal genes are required to maintain the cytoplasmic kappa particles.
Without a dominant gene this particle would disappear from the cytoplasm of the
host.![]()
![]()
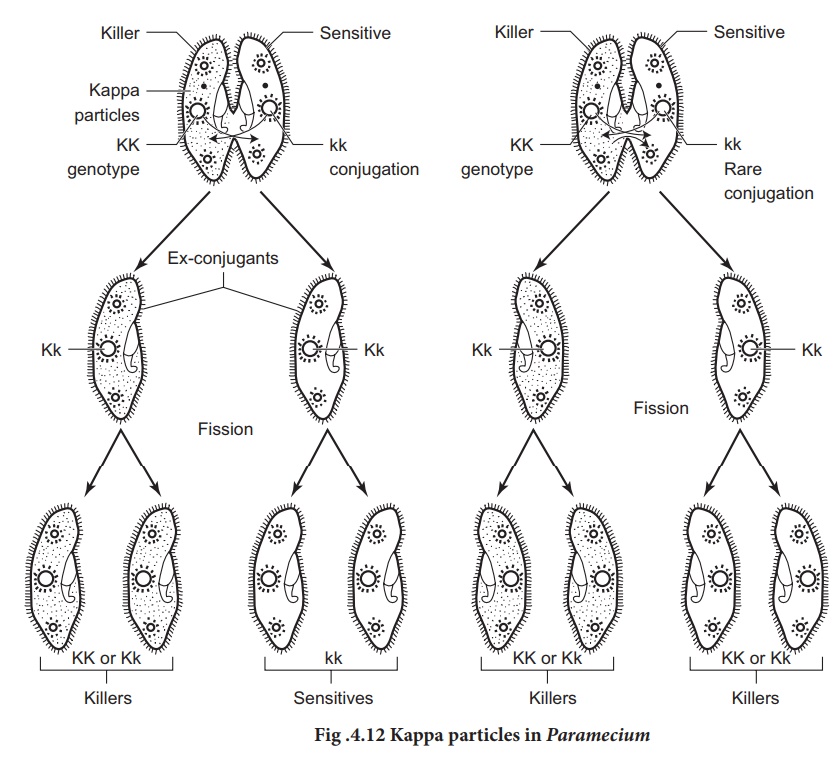
The kappa appears to be a bacterium, Caedobacter taeniospiralis that has its own DNA and replicates autonomously. Kappa particle occurs in atleast two forms; N and B forms. The N form is the infective form that passes from one Paremicium to another and confers the killer specificity to the host cell. The “N” form is attacked by a bacteriophage that induces the formation of inculsions called “R” bodies, inside the kappa particles and convert it to the “B” form.
These “R” bodies are visible under the light microscope as refractile
bodies. In the “B” form, kappa can no longer replicate, it is offen lysed
within the cell, however, it confers killer specificity on the host cell.
Whether viral DNA or kappa DNA codes the toxin paramecin is not known at
present.
Related Topics