Chapter: 11th Microbiology : Chapter 13 : Immunology
Organs of the Immune System
Organs of the Immune System
The immune system consists of structurally varied organs that
are distributed throughout the body. Based on function, the organs can be divided
into primary and secondary lymphoid organs (Figure 13.2). The primary lymphoid
organs are responsible for providing the appropriate microenvironments for the
development and maturation of antigen sensitive B and T cells. The thymus is
the primary lymphoid organ for development of T cells and the bone marrow is
the primary lymphoid organ for development of B cells. The secondary lymphoid
organs serve as sites where lymphocytes interact with antigen and undergo
proliferation and differentiation into antigen specific effector cells. The
spleen, lymph nodes and mucosal associated lymphoid tissues (MALT) are
secondary lymphoid organs. These are discussed in more detail below.
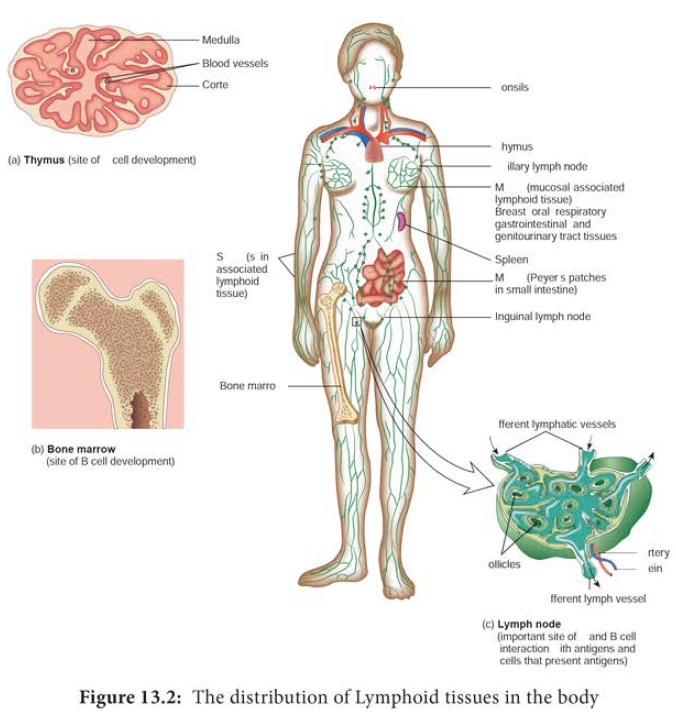
Primary Lymphoid Organs
a. Thymus
The thymus is a highly organized
lymphoid organ located above the heart. The thymus consists of two lobes.
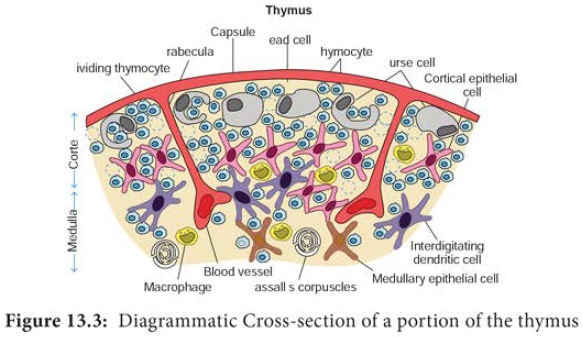
Each Lobe is surrounded by a capsule
and is divided into several lobules
by strands of connective tissue called trabeculae.
Each lobule contains an outer cortex and an inner medulla. The cortex contains many dividing immature lymphocytes and
isolated Hassall’s corpuscles (Figure 13.3). The primary function of the thymus
is the production of mature T cells. Precursor cells from the bone marrow
migrate into the outer cortex where they proliferate. As they mature, about 98%
die. This is due to a process known as thymic selection in which T cells that
recognize host (self) antigens are destroyed. The remaining 2% move into the
medulla of the thymus, become mature T cells and subsequently enter the blood
stream. These T cells recognize non host (non self) antigens.
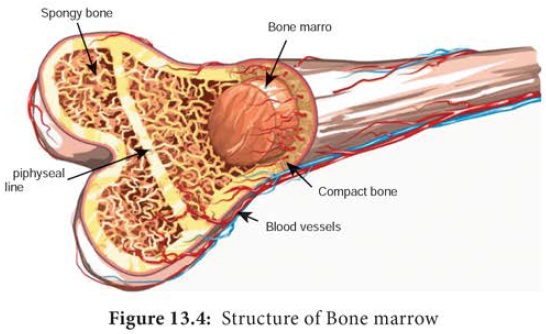
b. Bone marrow
In mammals, the bone
marrow (Figure 13.4) is the site of B cell maturation. Stromal cells within the bone marrow interact directly with the
Like thymic selection during T cell maturation, a selection
process within the bone marrow eliminates non functioning B cells and those
bearing self reactive antigen receptors. In birds, undifferentiated lymphocytes
move from the bone marrow to the Bursa
of Fabricius, where B cell mature; this is where B cells were first
identified and how they came to be known as “B” (for bursa) cells.
Secondary Lymphoid Organs
a. Spleen
The Spleen is the most highly organized secondary lymphoid organ. The spleen is a fist sized organ just behind the stomach. It collects and disposes of aged red blood cells. Its organization is shown schematically in Figure 13.5. The bulk of the spleen is composed of red pulb which is the site of red blood cell disposal. The spleen is not supplied by lymphatic vessels.
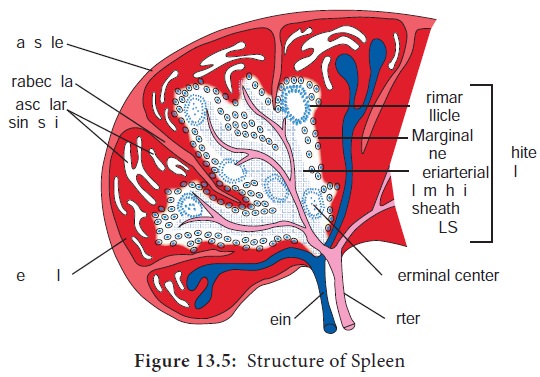
The lymphocytes surround the arterioles entering the spleen, forming areas of white pulp. The inner region of white pulp is divided into a Periarteriolar Lymphoid Sheath (PALS) containing mainly T cells. The spleen filters the blood and traps blood
borne microorganisms and antigens. Once trapped by splenic macrophages or
dendritic cells, the pathogen is phagocytosed, killed and digested.
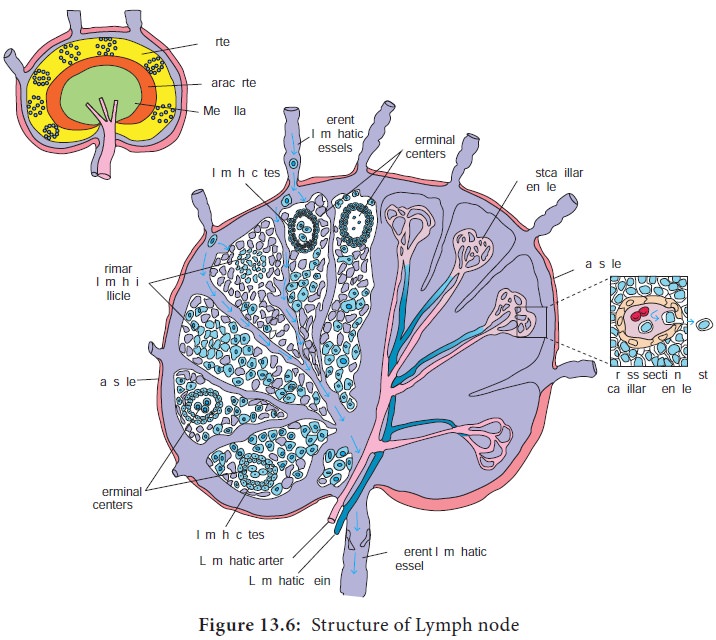
b. Lymph nodes
The lymph nodes are encapsulated
round structures located at the junction of major lymphatic vessels. Lymph node is
morphologically divided into three regions: the cortex, the paracortex and the
medulla (Figure 13.6). The outer most layer, the cortex contains lymphocytes
(mostly B cells), macrophages and follicular dendritic cells arranged in
primary follicles. After antigenic challenge, the primary follicles enlarge
into secondary follicles, each containing a germinal centre. Beneath the cortex
is the paracortex which is populated largely by T lymphocytes
and also interdigitating dendritic cells thought to have migrated from tissues
to the node. These interdigitating dentritic cells express high levels of class
II MHC molecules, which are necessary for presenting antigen to T helper (TH)
cells. Lymph nodes taken from neonatally thymectomized (removal of thymus from
new born animal) mice have unusually few cells in the paracortical region; the
paracortex is therefore sometimes referred to as a thymus dependent area in contrast to the cortex, which is a thymus independent area. The inner most layer of a lymph node, the medulla, is more sparsely populated with lymphoid lineage
cells; of those present many are plasma cells actively secreting antibody
molecules. Lymph nodes are specialized to trap antigen from regional tissue
spaces. As antigen is carried into a lymph node by the lymph, it is trapped,
processed and presented together with class II MHC molecules by interdigitating
dendritic cells in the paracortex, resulting
Activated TH
cells release cytokines needed for B cell activation. Thus lymph nodes
represent one environment where B cells differentiate into memory cells and
antibody – secreting plasma cells.
Lymph draining the extra cellular spaces of the body carries
antigen from the tissues to the lymph node through the afferent lymphatics. Lymph leaves by the efferent lymphatic in the medulla. Naive lymphocytes (mature lymphocytes not yet exposed to an antigen)
enter the node from the blood stream through specialized post capillary venules and leave with the lymph through the
efferent lymphatic.
c. MALT and SALT
The specialized lymphoid tissue in mucus membranes is called mucosal associated lymphoid tissue (MALT). There are
several types of MALT. The system most studied is the gut associated lymphoid tissue (GALT). GALT include the tonsils, adenoids, and appendix and specialized
structures called peyer’s patches (Figure13.7) in the
small intestine, which collect antigen from the epithelial surfaces of
the gastrointestinal tract. In peyer’s patches, the antigen is collected by
specialized epithelial cells called M cells (Figure 13.8). The
lymphocytes form a follicle consisting
of a large central dome of B lymphocytes surrounded by small number of T
lymphocytes. Similar but more diffusely organized aggregates of lymphocytes
protect the respiratory epithelium, where they are known as bronchial- associated lymphoid tissue (BALT).
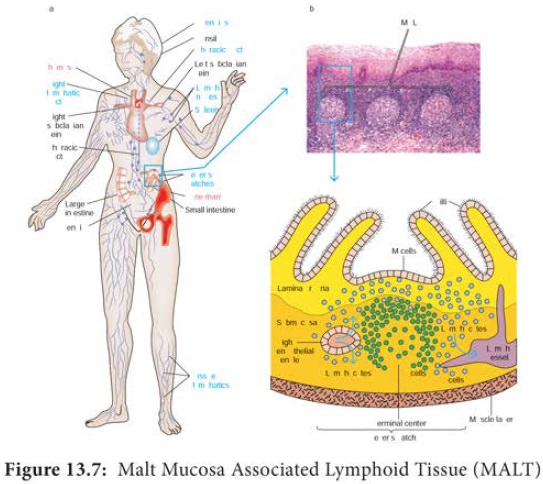
(a) The Peyer’s patch is a representative of the extensive MALT
system that is found in the instestine. (b) A stained tissue cross-section of
Peyer’s patch lymphoid nodules in the intestinal submucosa is schematically
diagrammed in (c). The intestinal lamina propria contains loose clusters of
lymphoid cells and diffuse follicles.
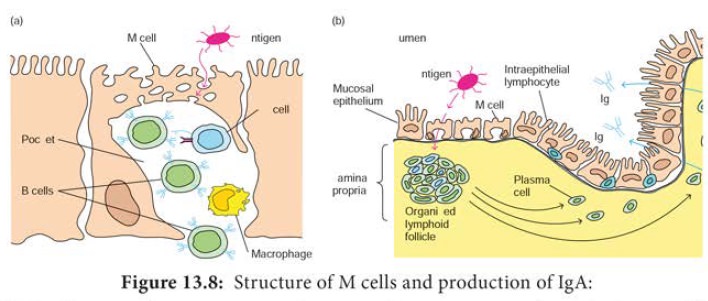
(a) M cells, situated in muscous membranes, endocytose antigen
from the lumen of the digestive, respiratory, and urogenitial tracts. The
antigen is transported across the cell and released into the large basolateral
pocket. (b) Antigen transported across the epithelial layer by M cells at an
inductive site activates B cells in the underlying lymphoid follicles. The
activated B cells differentiate into lgA-producing plasma cells, which migrate
along the lamina propria, the layer under the mucosa. The outer mucosal
epithelial layer contains intraepithelial lymphocytes, of which many are T
cells.
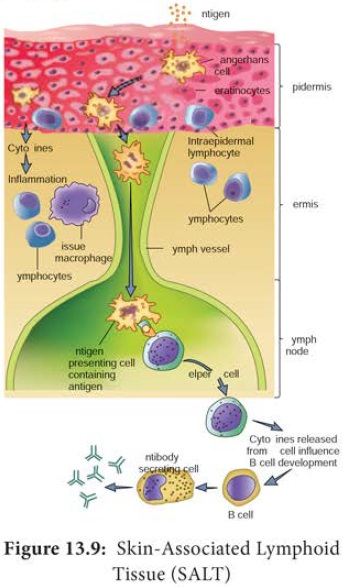
Despite the skin’s defenses, at times pathogenic microorganisms
gain access to the tissue under the skin surface. Here, they encounter a
specialized set of cells called the skin
associated lymphoid tissue (SALT) (Figure 13.9). The major function of SALT
is to confine microbial invaders to the area immediately underlying the
epidermis and to prevent them from gaining access to the blood stream. One type
of SALT is the langerhans cell, a
specialized myeloid cell that can phagocytose antigens.
Related Topics