Chapter: Medical Physiology: Introduction to Physiology: The Cell and Its Functions
Physical Structure of the Cell
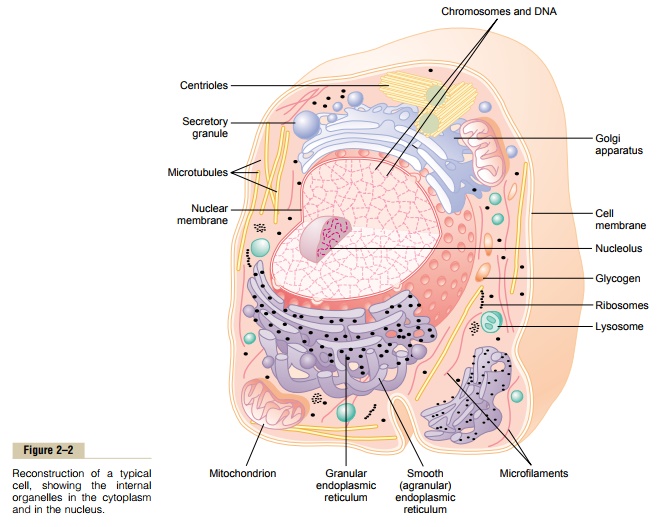
Physical Structure of the Cell
The cell is not merely a bag of fluid, enzymes, and chemicals; it also contains highly organized physical structures, called intracellular organelles. The physical nature of each organelle is as important as the cell’s chemical constituents for cell function. For instance, without one of the organelles, the mitochondria, more than 95 per cent of the cell’s energy release from nutri- ents would cease immediately. The most important organelles and other structures of the cell are shown in Figure 2–2.
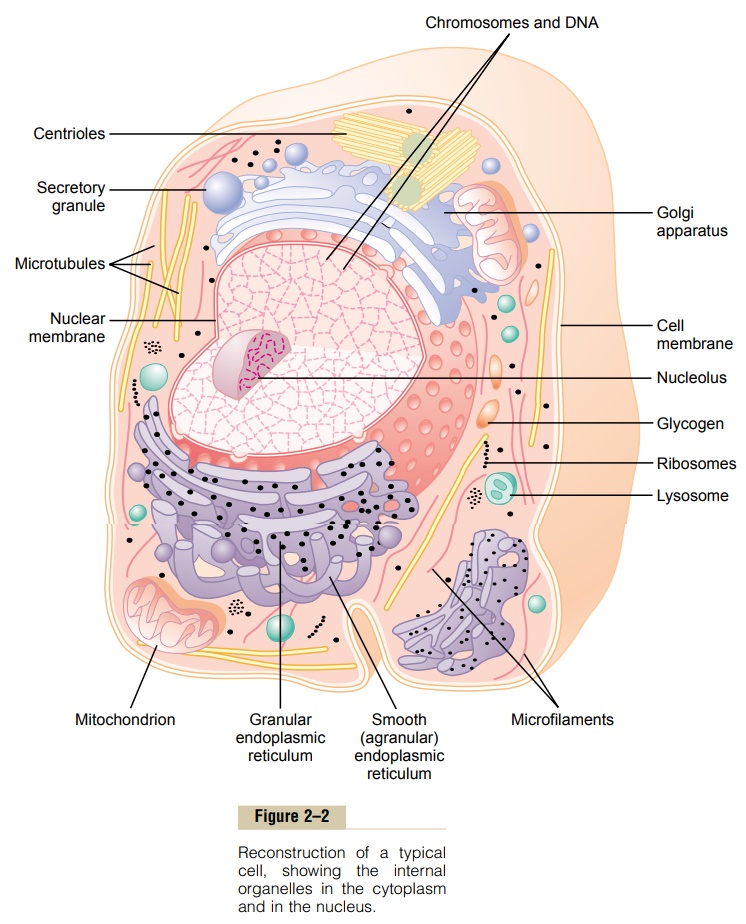
Membranous Structures of the Cell
Most organelles of the cell are covered by membranes composed primarily of lipids and proteins.These mem- branes include the cell membrane, nuclear membrane, membrane of the endoplasmic reticulum,and mem- branes of the mitochondria, lysosomes, and Golgi apparatus.
The lipids of the membranes provide a barrier that impedes the movement of water and water-soluble substances from one cell compartment to another because water is not soluble in lipids. However, protein molecules in the membrane often do penetrate all the way through the membrane, thus providing specialized pathways, often organized into actual pores, for passage of specific substances through the membrane.Also, many other membrane proteins are enzymes that catalyze a multitude of different chemical reactions, discussed here and in subsequent.
Cell Membrane
The cell membrane (also called the plasma mem- brane), which envelops the cell, is a thin, pliable, elastic structure only 7.5 to 10 nanometers thick. It is composed almost entirely of proteins and lipids.The approximate composition is proteins, 55 per cent; phospholipids, 25 per cent; cholesterol, 13 per cent; other lipids, 4 per cent; and carbohydrates, 3 per cent.
Lipid Barrier of the Cell Membrane Impedes Water Penetration.
Figure 2–3 shows the structure of the cell membrane. Its basic structure is a lipid bilayer, which is a thin, double-layered film of lipids—each layer only one molecule thick—that is continuous over the entire cell surface. Interspersed in this lipid film are large globu-lar protein molecules.
The basic lipid bilayer is composed of phospholipid molecules. One end of each phospholipid molecule is soluble in water; that is, it is hydrophilic. The other end is soluble only in fats; that is, it ishydrophobic. The
Because the hydrophobic portions of the phospho-lipid molecules are repelled by water but are mutually attracted to one another, they have a natural tendency to attach to one another in the middle of the mem-brane, as shown in Figure 2–3. The hydrophilic phos-phate portions then constitute the two surfaces of the complete cell membrane, in contact with intracellular water on the inside of the membrane andextracellular water on the outside surface.
The lipid layer in the middle of the membrane is impermeable to the usual water-soluble substances, such as ions, glucose, and urea. Conversely, fat-soluble substances, such as oxygen, carbon dioxide, and alcohol, can penetrate this portion of the membrane with ease.
The cholesterol molecules in the membrane are also lipid in nature because their steroid nucleus is highly fat soluble. These molecules, in a sense, are dissolved in the bilayer of the membrane. They mainly help determine the degree of permeability (or imperme-ability) of the bilayer to water-soluble constituents of body fluids. Cholesterol controls much of the fluidity of the membrane as well.
Cell Membrane Proteins. Figure 2–3 also shows globularmasses floating in the lipid bilayer. These are mem-brane proteins, most of which are glycoproteins. Two types of proteins occur: integral proteins that protrude all the way through the membrane, and peripheral pro-teins that are attached only to one surface of the mem-brane and do not penetrate all the way through.
Many of the integral proteins provide structural channels (or pores) through which water moleculesand water-soluble substances, especially ions, can diffuse between the extracellular and intracellular fluids. These protein channels also have selective prop-erties that allow preferential diffusion of some sub-stances over others.
Other integral proteins act as carrier proteins for transporting substances that otherwise could not pen-etrate the lipid bilayer. Sometimes these even trans-port substances in the direction opposite to their natural direction of diffusion, which is called “active transport.” Still others act as enzymes.
Integral membrane proteins can also serve as recep-tors for water-soluble chemicals, such as peptide hor-mones, that do not easily penetrate the cell membrane. Interaction of cell membrane receptors with specific ligands that bind to the receptor causes conforma-tional changes in the receptor protein. This, in turn, enzymatically activates the intracellular part of the protein or induces interactions between the receptor and proteins in the cytoplasm that act as second mes-sengers, thereby relaying the signal from the extracel-lular part of the receptor to the interior of the cell. In this way, integral proteins spanning the cell membrane provide a means of conveying information about the environment to the cell interior.
Peripheral protein molecules are often attached to the integral proteins. These peripheral proteins func-tion almost entirely as enzymes or as controllers of transport of substances through the cell membrane “pores.”
Membrane Carbohydrates—The Cell “Glycocalyx.” Mem-brane carbohydrates occur almost invariably in combination with proteins or lipids in the form of gly-coproteins or glycolipids. In fact, most of the integralproteins are glycoproteins, and about one tenth of the membrane lipid molecules are glycolipids. The “glyco” portions of these molecules almost invariably protrude to the outside of the cell, dangling outward from the cell surface. Many other carbohydrate compounds, called proteoglycans—which are mainly carbohydrate substances bound to small protein cores—are loosely attached to the outer surface of the cell as well. Thus, the entire outside surface of the cell often has a loose carbohydrate coat called the glycocalyx.
The carbohydrate moieties attached to the outer surface of the cell have several important functions:
(1) Many of them have a negative electrical charge, which gives most cells an overall negative surface charge that repels other negative objects. (2) The gly-cocalyx of some cells attaches to the glycocalyx of other cells, thus attaching cells to one another. (3) Many of the carbohydrates act as receptor substances for binding hormones, such as insulin; when bound, this combination activates attached internal proteins that, in turn, activate a cascade of intracellular enzymes. (4) Some carbohydrate moieties enter into immune reactions.
Cytoplasm and Its Organelles
The cytoplasm is filled with both minute and large dis-persed particles and organelles. The clear fluid portion of the cytoplasm in which the particles are dispersed is called cytosol; this contains mainly dissolved pro-teins, electrolytes, and glucose.
Dispersed in the cytoplasm are neutral fat globules, glycogen granules, ribosomes, secretory vesicles, and five especially important organelles: the endoplasmicreticulum, the Golgi apparatus, mitochondria, lyso-somes, and peroxisomes.
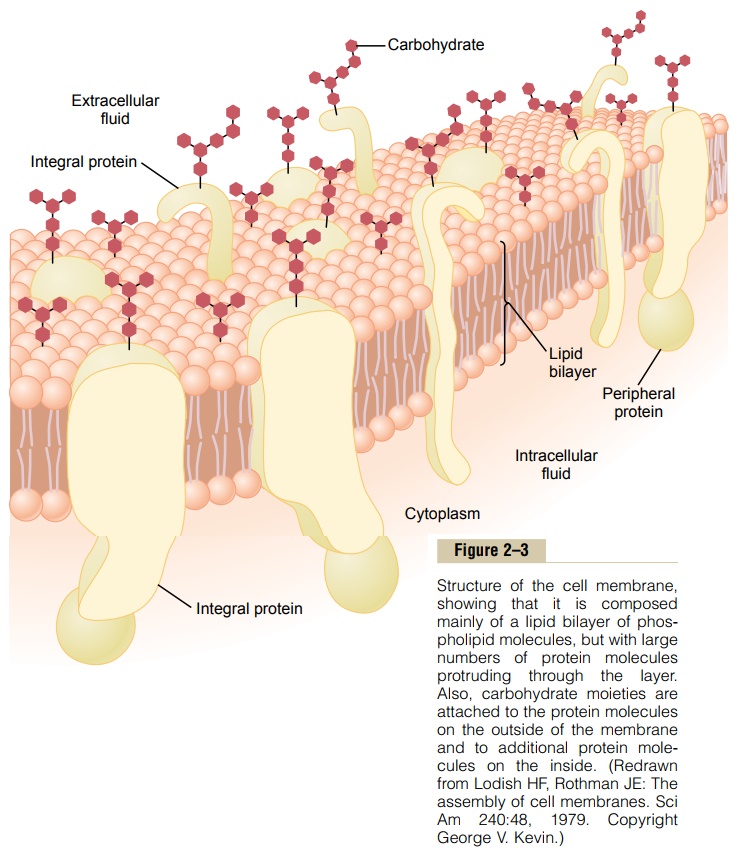
Endoplasmic Reticulum
Figure 2–2 shows a network of tubular and flat vesic-ular structures in the cytoplasm; this is the endoplas-mic reticulum. The tubules and vesicles interconnectwith one another. Also, their walls are constructed of lipid bilayer membranes that contain large amounts of proteins, similar to the cell membrane. The total surface area of this structure in some cells—the liver cells, for instance—can be as much as 30 to 40 times the cell membrane area.
The detailed structure of a small portion of endo-plasmic reticulum is shown in Figure 2–4. The space inside the tubules and vesicles is filled with endoplas-mic matrix, a watery medium that is different from thefluid in the cytosol outside the endoplasmic reticulum. Electron micrographs show that the space inside the endoplasmic reticulum is connected with the space between the two membrane surfaces of the nuclear membrane.
Substances formed in some parts of the cell enter the space of the endoplasmic reticulum and are then conducted to other parts of the cell. Also, the vast surface area of this reticulum and the multiple enzyme systems attached to its membranes provide machinery for a major share of the metabolic functions of the cell.
Ribosomes and the Granular Endoplasmic Reticulum.
Attached to the outer surfaces of many parts of the endoplasmic reticulum are large numbers of minute granular particles called ribosomes.
Where these are present, the reticulum is called the granular endoplas-mic reticulum. The ribosomes are composed of amixture of RNA and proteins, and they function to synthesize new protein molecules in the cell.
Agranular Endoplasmic Reticulum. Part of the endoplasmicreticulum has no attached ribosomes. This part is called the agranular, or smooth, endoplasmic reticu-lum. The agranular reticulum functions for the syn-thesis of lipid substances and for other processes of the cells promoted by intrareticular enzymes.
Golgi Apparatus
The Golgi apparatus, shown in Figure 2–5, is closely related to the endoplasmic reticulum. It has mem-branes similar to those of the agranular endoplasmic reticulum. It usually is composed of four or more stacked layers of thin, flat, enclosed vesicles lying near one side of the nucleus. This apparatus is prominent in secretory cells, where it is located on the side of the cell from which the secretory substances are extruded.
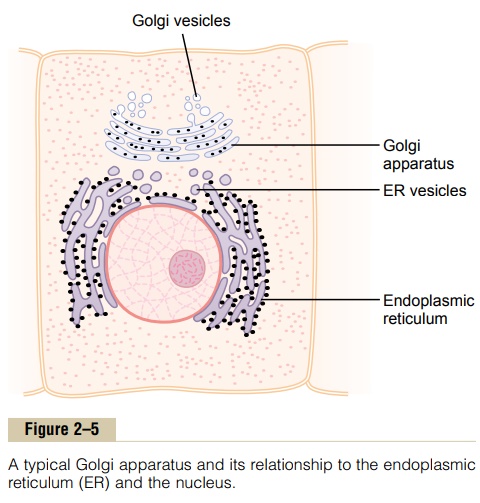
The Golgi apparatus functions in association with the endoplasmic reticulum. As shown in Figure 2–5, small “transport vesicles” (also called endoplasmic reticulum vesicles, or ER vesicles) continually pinch off from the endoplasmic reticulum and shortly thereafter fuse with the Golgi apparatus. In this way, substances entrapped in the ER vesicles are transported from the endoplasmic reticulum to the Golgi apparatus. The transported substances are then processed in the Golgi apparatus to form lysosomes, secretory vesicles, and other cytoplasmic components.
Lysosomes
Lysosomes, shown in Figure 2–2, are vesicular organelles that form by breaking off from the Golgi apparatus and then dispersing throughout the cyto-plasm. The lysosomes provide an intracellular digestivesystem that allows the cell to digest (1) damaged cel-lular structures, (2) food particles that have been ingested by the cell, and (3) unwanted matter such as bacteria. The lysosome is quite different in different types of cells, but it is usually 250 to 750 nanometers in diameter. It is surrounded by a typical lipid bilayer membrane and is filled with large numbers of small granules 5 to 8 nanometers in diameter, which are protein aggregates of as many as 40 different hydro-lase (digestive) enzymes. A hydrolytic enzyme iscapable of splitting an organic compound into two or more parts by combining hydrogen from a water mol-ecule with one part of the compound and combining the hydroxyl portion of the water molecule with the other part of the compound. For instance, protein is hydrolyzed to form amino acids, glycogen is hydrolyzed to form glucose, and lipids are hydrolyzed to form fatty acids and glycerol.
Ordinarily, the membrane surrounding the lysosome prevents the enclosed hydrolytic enzymes from coming in contact with other substances in the cell and, therefore, prevents their digestive actions. However, some conditions of the cell break the membranes of some of the lysosomes, allowing release of the diges-tive enzymes. These enzymes then split the organic substances with which they come in contact into small, highly diffusible substances such as amino acids and glucose.
Peroxisomes
Peroxisomes are similar physically to lysosomes, but they are different in two important ways. First, they are believed to be formed by self-replication (or perhaps by budding off from the smooth endoplasmic reticu-lum) rather than from the Golgi apparatus. Second, they contain oxidases rather than hydrolases. Several of the oxidases are capable of combining oxygen with hydrogen ions derived from different intracellular chemicals to form hydrogen peroxide (H2O2). Hydro-gen peroxide is a highly oxidizing substance and is used in association with catalase, another oxidase enzyme present in large quantities in peroxisomes, to oxidize many substances that might otherwise be poi-sonous to the cell. For instance, about half the alcohol a person drinks is detoxified by the peroxisomes of the liver cells in this manner.
Secretory Vesicles
One of the important functions of many cells is secre-tion of special chemical substances. Almost all such secretory substances are formed by the endoplasmic reticulum–Golgi apparatus system and are then released from the Golgi apparatus into the cytoplasm in the form of storage vesicles called secretory vesicles or secretory granules. Figure 2–6 shows typical secre-tory vesicles inside pancreatic acinar cells; these
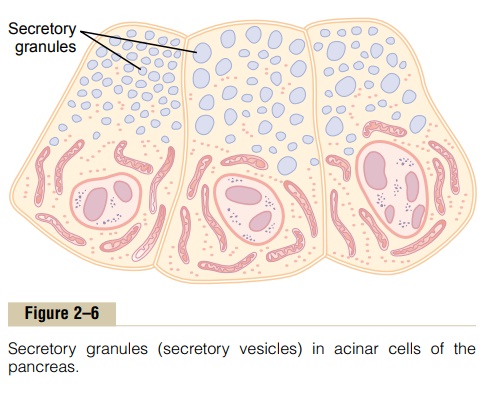
vesicles store protein proenzymes (enzymes that are not yet activated). The proenzymes are secreted later through the outer cell membrane into the pancreatic duct and thence into the duodenum, where they become activated and perform digestive functions on the food in the intestinal tract.
Mitochondria
The mitochondria, shown in Figures 2–2 and 2–7, are called the “powerhouses” of the cell. Without them, cells would be unable to extract enough energy from the nutrients, and essentially all cellular functions would cease.
Mitochondria are present in all areas of each cell’s cytoplasm, but the total number per cell varies from less than a hundred up to several thousand, depending on the amount of energy required by the cell. Further, the mitochondria are concentrated in those portions of the cell that are responsible for the major share of its energy metabolism. They are also variable in size and shape. Some are only a few hundred nanometers in diameter and globular in shape, whereas others are elongated—as large as 1 micrometer in diameter and 7 micrometers long; still others are branching and filamentous.
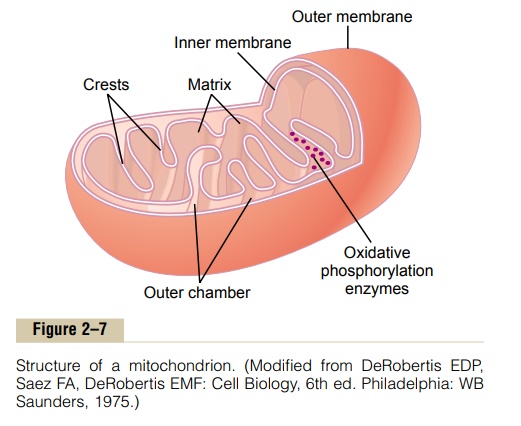
The basic structure of the mitochondrion, shown in Figure 2–7, is composed mainly of two lipid bilayer–protein membranes: an outer membrane and an inner membrane. Many infoldings of the inner membrane form shelves onto which oxidative enzymes are attached. In addition, the inner cavity of the mito-chondrion is filled with a matrix that contains large quantities of dissolved enzymes that are necessary for extracting energy from nutrients. These enzymes operate in association with the oxidative enzymes on the shelves to cause oxidation of the nutrients, thereby forming carbon dioxide and water and at the same time releasing energy. The liberated energy is used to synthesize a “high-energy” substance called adenosinetriphosphate (ATP). ATP is then transported out of themitochondrion, and it diffuses throughout the cell to release its own energy wherever it is needed for per-forming cellular functions.
Mitochondria are self-replicative, which means that one mitochondrion can form a second one, a third one, and so on, whenever there is a need in the cell for increased amounts of ATP. Indeed, the mitochondria contain DNA similar to that found in the cell nucleus. We will see that DNA is the basic chem-ical of the nucleus that controls replication of the cell. The DNA of the mitochondrion plays a similar role, controlling replication of the mitochondrion itself.
Filament and Tubular Structures of the Cell
The fibrillar proteins of the cell are usually organized into filaments or tubules. These originate as precursor protein molecules synthesized by ribosomes in the cytoplasm. The precursor molecules then polymerize to form filaments. As an example, large numbers of actin filaments frequently occur in the outer zone of the cytoplasm, called the ectoplasm, to form an elastic support for the cell membrane. Also, in muscle cells, actin and myosin filaments are organized into a special contractile machine that is the basis for muscle con-traction.
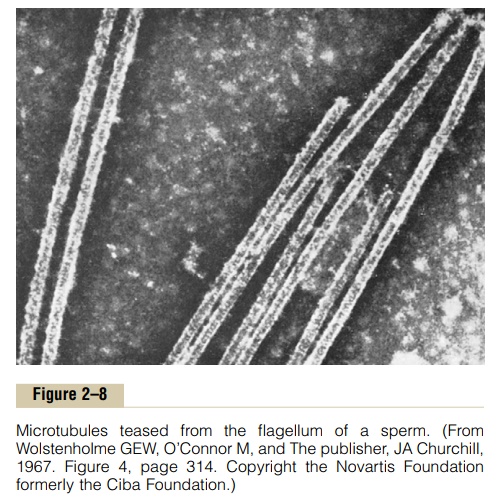
A special type of stiff filament composed of poly-merized tubulin molecules is used in all cells to con-struct very strong tubular structures, the microtubules. Figure 2–8 shows typical microtubules that were teased from the flagellum of a sperm.
Another example of microtubules is the tubular skeletal structure in the center of each cilium that radi-ates upward from the cell cytoplasm to the tip of the cilium. This structure is discussed later and is illustrated in Figure 2–17. Also, both the centri-oles and the mitotic spindle of the mitosing cell arecomposed of stiff microtubules.
Thus, a primary function of microtubules is to act as a cytoskeleton, providing rigid physical structures for certain parts of cells.
Nucleus
The nucleus is the control center of the cell. Briefly, the nucleus contains large quantities of DNA, which are the genes. The genes determine the characteristics of the cell’s proteins, including the structural proteins, as well as the intracellular enzymes that control cyto-plasmic and nuclear activities.
The genes also control and promote reproduction of the cell itself. The genes first reproduce to give two identical sets of genes; then the cell splits by a special process called mitosis to form two daughter cells, each of which receives one of the two sets of DNA genes.
Unfortunately, the appearance of the nucleus under the microscope does not provide many clues to the mechanisms by which the nucleus performs its control activities. Figure 2–9 shows the light microscopic appearance of the interphase nucleus (during the period between mitoses), revealing darkly staining chromatin material throughout the nucleoplasm.During mitosis, the chromatin material organizes in the form of highly structured chromosomes, which can then be easily identified using the light microscope.
Nuclear Membrane
The nuclear membrane, also called the nuclear enve-lope, is actually two separate bilayer membranes, oneinside the other. The outer membrane is continuous with the endoplasmic reticulum of the cell cytoplasm, and the space between the two nuclear membranes is also continuous with the space inside the endoplasmic reticulum, as shown in Figure 2–9.
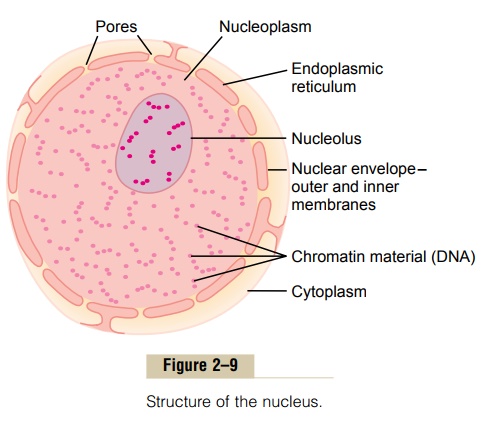
The nuclear membrane is penetrated by several thousand nuclear pores. Large complexes of protein molecules are attached at the edges of the pores so that the central area of each pore is only about 9 nanometers in diameter. Even this size is large enough to allow molecules up to 44,000 molecular weight to pass through with reasonable ease.
Nucleoli and Formation of Ribosomes
The nuclei of most cells contain one or more highly staining structures called nucleoli. The nucleolus, unlike most other organelles discussed here, does not have a limiting membrane. Instead, it is simply an accu-mulation of large amounts of RNA and proteins of the types found in ribosomes. The nucleolus becomes con-siderably enlarged when the cell is actively synthesiz-ing proteins.
Formation of the nucleoli (and of the ribosomes in the cytoplasm outside the nucleus) begins in the nucleus. First, specific DNA genes in the chromosomes cause RNA to be synthesized. Some of this is stored in the nucleoli, but most of it is transported outward through the nuclear pores into cytoplasm. Here, it is used in conjunction with specific proteins to assemble “mature” ribosomes that play an essential role in forming cytoplasmic proteins.
Related Topics