Chapter: Medical Physiology: Introduction to Physiology: The Cell and Its Functions
Functional Systems of the Cell
Functional Systems of the Cell
Ingestion by the Cell—Endocytosis
If a cell is to live and grow and reproduce, it must obtain nutrients and other substances from the sur-rounding fluids. Most substances pass through the cell membrane by diffusion and active transport.
Diffusion involves simple movement through the membrane caused by the random motion of the mol-ecules of the substance; substances move either through cell membrane pores or, in the case of lipid-soluble substances, through the lipid matrix of the membrane.
Active transport involves the actual carrying of a substance through the membrane by a physical pro-tein structure that penetrates all the way through the membrane.
Very large particles enter the cell by a specialized function of the cell membrane called endocytosis. The principal forms of endocytosis are pinocytosis and phagocytosis. Pinocytosis means ingestion of minuteparticles that form vesicles of extracellular fluid and particulate constituents inside the cell cytoplasm. Phagocytosis means ingestion of large particles, such as bacteria, whole cells, or portions of degenerating tissue.
Pinocytosis. Pinocytosis occurs continually in the cellmembranes of most cells, but it is especially rapid in some cells. For instance, it occurs so rapidly in macrophages that about 3 per cent of the total macro-phage membrane is engulfed in the form of vesicles each minute. Even so, the pinocytotic vesicles are so small—usually only 100 to 200 nanometers in diame-ter—that most of them can be seen only with the elec-tron microscope.
Pinocytosis is the only means by which most large macromolecules, such as most protein molecules, can enter cells. In fact, the rate at which pinocytotic vesi-cles form is usually enhanced when such macro-molecules attach to the cell membrane.
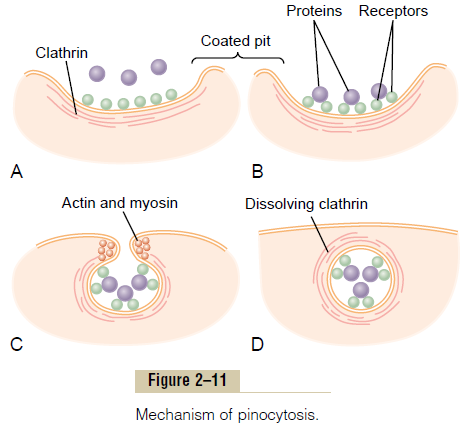
Figure 2–11 demonstrates the successive steps of pinocytosis, showing three molecules of protein attaching to the membrane. These molecules usually attach to specialized protein receptors on the surface of the membrane that are specific for the type of protein that is to be absorbed. The receptors generally are concentrated in small pits on the outer surface of the cell membrane, called coated pits. On the inside of the cell membrane beneath these pits is a latticework of fibrillar protein called clathrin, as well as other pro-teins, perhaps including contractile filaments of actin and myosin.
Once the protein molecules have bound with the receptors, the surface properties of the local membrane change in such a way that the entire pit invaginates inward, and the fibrillar proteins sur-rounding the invaginating pit cause its borders to close over the attached proteins as well as over a small amount of extracellular fluid. Immediately thereafter, the invaginated portion of the membrane breaks away from the surface of the cell, forming a pinocytoticvesicle inside the cytoplasm of the cell.
What causes the cell membrane to go through the necessary contortions to form pinocytotic vesicles remains mainly a mystery.This process requires energy from within the cell; this is supplied by ATP, a high-energy substance discussed later. Also, it requires the presence of calcium ions in the extra-cellular fluid, which probably react with contractile protein filaments beneath the coated pits to provide the force for pinching the vesicles away from the cell membrane.
Phagocytosis. Phagocytosis occurs in much the sameway as pinocytosis, except that it involves large particles rather than molecules. Only certain cells have the capability of phagocytosis, most notably the tissue macrophages and some of the white blood cells.
Phagocytosis is initiated when a particle such as a bacterium, a dead cell, or tissue debris binds with receptors on the surface of the phagocyte. In the case of bacteria, each bacterium usually is already attached to a specific antibody, and it is the antibody that attaches to the phagocyte receptors, dragging the bac-terium along with it. This intermediation of antibodies is called opsonization.
Phagocytosis occurs in the following steps:
1. The cell membrane receptors attach to the surface ligands of the particle.
2. The edges of the membrane around the points of attachment evaginate outward within a fraction of a second to surround the entire particle; then, progressively more and more membrane receptors attach to the particle ligands. All this occurs suddenly in a zipper-like manner to form a closed phagocytic vesicle.
3. Actin and other contractile fibrils in the cytoplasm surround the phagocytic vesicle and contract around its outer edge, pushing the vesicle to the interior.
4. The contractile proteins then pinch the stem of the vesicle so completely that the vesicle separates from the cell membrane, leaving the vesicle in the cell interior in the same way that pinocytotic vesicles are formed.
Digestion of Pinocytotic and Phagocytic Foreign Substances Inside the Cell—Function of the Lysosomes
Almost immediately after a pinocytotic or phagocytic vesicle appears inside a cell, one or more lysosomes become attached to the vesicle and empty their acidhydrolases to the inside of the vesicle, as shown in Figure 2–12. Thus, a digestive vesicle is formed inside the cell cytoplasm in which the vesicular hydrolases begin hydrolyzing the proteins, carbohydrates, lipids, and other substances in the vesicle. The products of digestion are small molecules of amino acids, glucose, phosphates, and so forth that can diffuse through the membrane of the vesicle into the cytoplasm. What is left of the digestive vesicle, called the residual body, represents indigestible substances. In most instances, this is finally excreted through the cell membrane by a process called exocytosis, which is essentially the opposite of endocytosis.
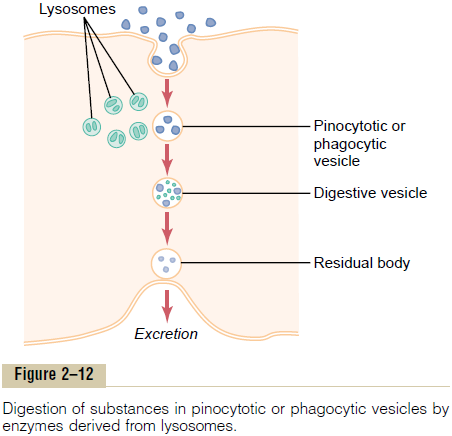
Thus, the pinocytotic and phagocytic vesicles con-taining lysosomes can be called the digestive organs of the cells.
Regression of Tissues and Autolysis of Cells. Tissues of thebody often regress to a smaller size. For instance, this occurs in the uterus after pregnancy, in muscles during long periods of inactivity, and in mammary glands at the end of lactation. Lysosomes are responsible for much of this regression. The mechanism by which lack of activity in a tissue causes the lysosomes to increase their activity is unknown.
Another special role of the lysosomes is removal of damaged cells or damaged portions of cells from tissues. Damage to the cell—caused by heat, cold, trauma, chemicals, or any other factor—induces lyso-somes to rupture. The released hydrolases immedi-ately begin to digest the surrounding organic substances. If the damage is slight, only a portion of the cell is removed, followed by repair of the cell. If the damage is severe, the entire cell is digested, a process called autolysis. In this way, the cell is com-pletely removed, and a new cell of the same type ordi-narily is formed by mitotic reproduction of an adjacent cell to take the place of the old one.
The lysosomes also contain bactericidal agents that can kill phagocytized bacteria before they can cause cellular damage. These agents include (1) lysozyme, which dissolves the bacterial cell membrane; (2)lyso-ferrin, which binds iron and other substances beforethey can promote bacterial growth; and (3) acid at a pH of about 5.0, which activates the hydrolases and inactivates bacterial metabolic systems.
Synthesis and Formation of Cellular Structures by Endoplasmic Reticulum and Golgi Apparatus
Specific Functions of the Endoplasmic Reticulum
The extensiveness of the endoplasmic reticulum and the Golgi apparatus in secretory cells has already been emphasized. These structures are formed primarily of lipid bilayer membranes similar to the cell membrane, and their walls are loaded with protein enzymes that catalyze the synthesis of many substances required by the cell.
Most synthesis begins in the endoplasmic reticulum. The products formed there are then passed on to the Golgi apparatus, where they are further processed before being released into the cytoplasm. But first, let us note the specific products that are synthesized in specific portions of the endoplasmic reticulum and the Golgi apparatus.
Proteins Are Formed by the Granular Endoplasmic Reticulum.
The granular portion of the endoplasmic reticulum is characterized by large numbers of ribosomes attached to the outer surfaces of the endoplasmic reticulum membrane. As we discuss, protein mole-cules are synthesized within the structures of the ribo-somes. The ribosomes extrude some of the synthesized protein molecules directly into the cytosol, but they also extrude many more through the wall of the endo-plasmic reticulum to the interior of the endoplasmic vesicles and tubules, that is, into the endoplasmicmatrix.
Synthesis of Lipids by the Smooth Endoplasmic Reticulum.
The endoplasmic reticulum also synthesizes lipids, especially phospholipids and cholesterol. These are rapidly incorporated into the lipid bilayer of the endo-plasmic reticulum itself, thus causing the endoplasmic reticulum to grow more extensive. This occurs mainly in the smooth portion of the endoplasmic reticulum.
To keep the endoplasmic reticulum from growing beyond the needs of the cell, small vesicles called ERvesicles or transport vesicles continually break awayfrom the smooth reticulum; most of these vesicles then migrate rapidly to the Golgi apparatus.
Other Functions of the Endoplasmic Reticulum. Other sig-nificant functions of the endoplasmic reticulum, espe-cially the smooth reticulum, include the following:
1. It provides the enzymes that control glycogen breakdown when glycogen is to be used for energy.
2. It provides a vast number of enzymes that are capable of detoxifying substances, such as drugs, that might damage the cell. It achieves detoxification by coagulation, oxidation, hydrolysis, conjugation with glycuronic acid, and in other ways.
Specific Functions of the Golgi Apparatus Synthetic Functions of the Golgi Apparatus. Although themajor function of the Golgi apparatus is to provide additional processing of substances already formed in the endoplasmic reticulum, it also has the capability of synthesizing certain carbohydrates that cannot be formed in the endoplasmic reticulum. This is especially true for the formation of large saccharide polymers bound with small amounts of protein; the most impor-tant of these are hyaluronic acid and chondroitin sulfate.
A few of the many functions of hyaluronic acid and chondroitin sulfate in the body are as follows: (1) they are the major components of proteoglycans secreted in mucus and other glandular secretions; (2) they are the major components of the ground substance outside the cells in the interstitial spaces, acting as filler between collagen fibers and cells; and (3) they are principal components of the organic matrix in both cartilage and bone.
Processing of Endoplasmic Secretions by the Golgi Apparatus— Formation of Vesicles. Figure 2–13 summarizes themajor functions of the endoplasmic reticulum and Golgi apparatus. As substances are formed in the endoplasmic reticulum, especially the proteins, they are transported through the tubules toward portions of the smooth endoplasmic reticulum that lie nearest the Golgi apparatus. At this point, small transport vesi-cles composed of small envelopes of smooth endo-plasmic reticulum continually break away and diffuse to the deepest layer of the Golgi apparatus. Inside these vesicles are the synthesized proteins and other prod-ucts from the endoplasmic reticulum.
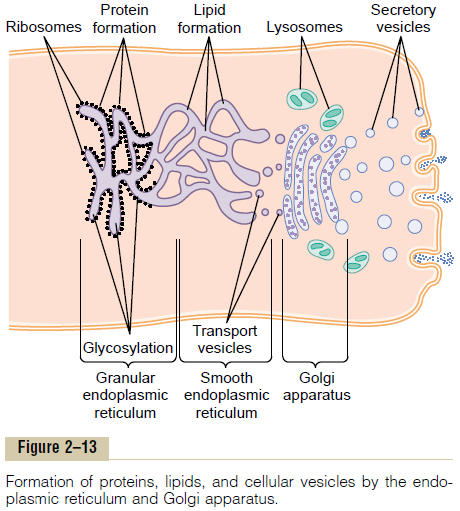
The transport vesicles instantly fuse with the Golgi apparatus and empty their contained substances into the vesicular spaces of the Golgi apparatus. Here, additional carbohydrate moieties are added to the secretions. Also, an important function of the Golgi apparatus is to compact the endoplasmic reticular secretions into highly concentrated packets. As the secretions pass toward the outermost layers of the Golgi apparatus, the compaction and processing proceed. Finally, both small and large vesicles contin-ually break away from the Golgi apparatus, carrying with them the compacted secretory substances, and in turn, the vesicles diffuse throughout the cell.
To give an idea of the timing of these processes: When a glandular cell is bathed in radioactive amino acids, newly formed radioactive protein molecules can be detected in the granular endoplasmic reticulum within 3 to 5 minutes. Within 20 minutes, newly formed proteins are already present in the Golgi apparatus, and within 1 to 2 hours, radioactive proteins are secreted from the surface of the cell.
Types of Vesicles Formed by the Golgi Apparatus—Secretory Vesicles and Lysosomes. In a highly secretory cell, thevesicles formed by the Golgi apparatus are mainly secretory vesicles containing protein substances thatare to be secreted through the surface of the cell mem-brane. These secretory vesicles first diffuse to the cell membrane, then fuse with it and empty their sub-stances to the exterior by the mechanism called exo-cytosis. Exocytosis, in most cases, is stimulated by theentry of calcium ions into the cell; calcium ions inter-act with the vesicular membrane in some way that is not understood and cause its fusion with the cell mem-brane, followed by exocytosis—that is, opening of the membrane’s outer surface and extrusion of its contents outside the cell.
Some vesicles, however, are destined for intracellu-lar use.
Use of Intracellular Vesicles to Replenish Cellular Membranes.
Some of the intracellular vesicles formed by the Golgi apparatus fuse with the cell membrane or with the membranes of intracellular structures such as the mitochondria and even the endoplasmic reticulum. This increases the expanse of these membranes and thereby replenishes the membranes as they are used up. For instance, the cell membrane loses much of its substance every time it forms a phagocytic or pinocy-totic vesicle, and the vesicular membranes of the Golgi apparatus continually replenish the cell membrane.
In summary, the membranous system of the endo-plasmic reticulum and Golgi apparatus represents a highly metabolic organ capable of forming new intra-cellular structures as well as secretory substances to be extruded from the cell.
Extraction of Energy from Nutrients— Function of the Mitochondria
The principal substances from which cells extract energy are foodstuffs that react chemically with oxygen—carbohydrates, fats, and proteins. In the human body, essentially all carbohydrates are con-verted into glucose by the digestive tract and liver before they reach the other cells of the body. Similarly, proteins are converted into amino acids and fats into fatty acids. Figure 2–14 shows oxygen and the food-stuffs—glucose, fatty acids, and amino acids—all enter-ing the cell. Inside the cell, the foodstuffs react chemically with oxygen, under the influence of enzymes that control the reactions and channel the energy released in the proper direction.
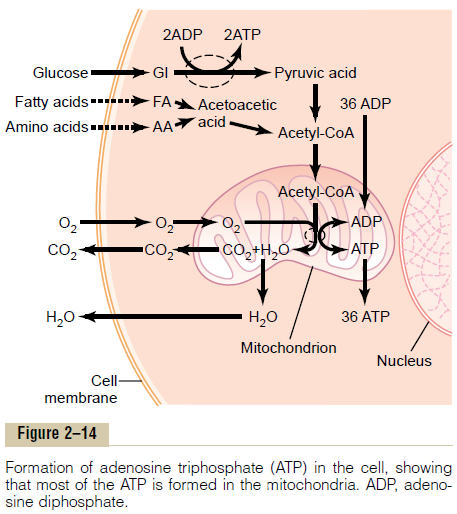
Briefly, almost all these oxidative reactions occur inside the mitochondria, and the energy that is released is used to form the high-energy compound ATP. Then, ATP, not the original foodstuffs, is usedthroughout the cell to energize almost all the subse-quent intracellular metabolic reactions.
Functional Characteristics of ATP
ATP is a nucleotide composed of (1) the nitrogenous base adenine, (2) the pentose sugar ribose, and (3) three phosphate radicals. The last two phosphate rad-icals are connected with the remainder of the molecule by so-called high-energy phosphate bonds, which are represented in the formula above by the symbol ~.
Under the physical and chemical conditions of the body, each of these high-energy bonds contains about12,000 calories of energy per mole of ATP, which is many times greater than the energy stored in the average chemical bond, thus giving rise to the term high-energy bond. Further, the high-energy phosphatebond is very labile, so that it can be split instantly on demand whenever energy is required to promote other intracellular reactions.
When ATP releases its energy, a phosphoric acid radical is split away, and adenosine diphosphate (ADP) is formed. This released energy is used to energize vir-tually all of the cell’s other functions, such as synthe-sis of substances and muscular contraction.
To reconstitute the cellular ATP as it is used up, energy derived from the cellular nutrients causes ADP and phosphoric acid to recombine to form new ATP, and the entire process repeats over and over again. For these reasons,ATP has been called the energy currency of the cell because it can be spent and remade contin-ually, having a turnover time of only a few minutes.
Chemical Processes in the Formation of ATP—Role of the Mitochondria. On entry into the cells, glucose is sub-jected to enzymes in the cytoplasm that convert it into pyruvic acid (a process called glycolysis). A small amount of ADP is changed into ATP by the energy released during this conversion, but this amount accounts for less than 5 per cent of the overall energy metabolism of the cell.
By far, the major portion of the ATP formed in the cell, about 95 per cent, is formed in the mitochondria. The pyruvic acid derived from carbohydrates, fatty acids from lipids, and amino acids from proteins are eventually converted into the compound acetyl-CoA in the matrix of the mitochondrion. This substance, in turn, is further dissoluted (for the purpose of extract-ing its energy) by another series of enzymes in the mitochondrion matrix, undergoing dissolution in a sequence of chemical reactions called the citric acidcycle, or Krebs cycle.
In this citric acid cycle, acetyl-CoA is split into its component parts, hydrogen atoms and carbon dioxide.
The carbon dioxide diffuses out of the mitochondria and eventually out of the cell; finally, it is excreted from the body through the lungs.
The hydrogen atoms, conversely, are highly reactive, and they combine instantly with oxygen that has also diffused into the mitochondria. This releases a tremen-dous amount of energy, which is used by the mito-chondria to convert very large amounts of ADP to ATP. The processes of these reactions are complex, requiring the participation of large numbers of protein enzymes that are integral parts of mitochondrial mem-branous shelves that protrude into the mitochondrialmatrix. The initial event is removal of an electron from the hydrogen atom, thus converting it to a hydrogen ion. The terminal event is combination of hydrogen ions with oxygen to form water plus the release of tremendous amounts of energy to large globular pro-teins, called ATP synthetase, that protrude like knobs from the membranes of the mitochondrial shelves. Finally, the enzyme ATP synthetase uses the energy from the hydrogen ions to cause the conversion of ADP to ATP. The newly formed ATP is transported out of the mitochondria into all parts of the cell cyto-plasm and nucleoplasm, where its energy is used to energize multiple cell functions.
This overall process for formation of ATP is called the chemiosmotic mechanism of ATP formation.
Uses of ATP for Cellular Function. Energy from ATP isused to promote three major categories of cellular functions: (1) transport of substances through multiple membranes in the cell, (2) synthesis of chemicalcompounds throughout the cell, and (3) mechanical work. These uses of ATP are illustrated by examplesin Figure 2–15: (1) to supply energy for the transport of sodium through the cell membrane, (2) to promote protein synthesis by the ribosomes, and (3) to supply the energy needed during muscle contraction.
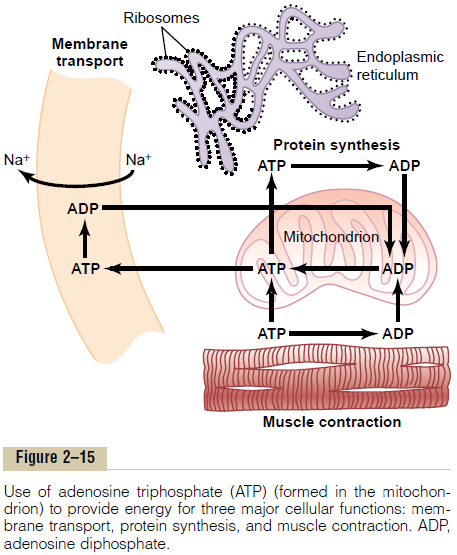
In addition to membrane transport of sodium, energy from ATP is required for membrane transport of potassium ions, calcium ions, magnesium ions, phosphate ions, chloride ions, urate ions, hydrogen ions, and many other ions and various organic substances. Membrane transport is so important to cell function that some cells—the renal tubular cells, for instance— use as much as 80 per cent of the ATP that they form for this purpose alone.
In addition to synthesizing proteins, cells synthesize phospholipids, cholesterol, purines, pyrimidines, and a host of other substances. Synthesis of almost any chemical compound requires energy. For instance, a single protein molecule might be composed of as many as several thousand amino acids attached to one another by peptide linkages; the formation of each of these linkages requires energy derived from the break-down of four high-energy bonds; thus, many thousand ATP molecules must release their energy as each protein molecule is formed. Indeed, some cells use as much as 75 per cent of all the ATP formed in the cell simply to synthesize new chemical compounds, espe-cially protein molecules; this is particularly true during the growth phase of cells.
The final major use of ATP is to supply energy for special cells to perform mechanical work. We see that each contraction of a muscle fiber requires expenditure of tremendous quantities of ATP energy. Other cells perform mechanical work in other ways, especially by ciliary and ameboid motion. The source of energy for all these types of mechanical work is ATP.
In summary, ATP is always available to release its energy rapidly and almost explosively wherever in the cell it is needed. To replace the ATP used by the cell,much slower chemical reactions break down carbohy-drates, fats, and proteins and use the energy derived from these to form new ATP. More than 95 per cent of this ATP is formed in the mitochondria, which accounts for the mitochondria being called the “pow-erhouses” of the cell.
Related Topics