Chapter: Plant Biochemistry: The use of energy from sunlight by photosynthesis is the basis of life on earth
How is the excitation energy of the photons captured in the antennae and transferred to the reaction centers?
How is the excitation energy of the photons captured in the antennae and transferred to the reaction centers?
The transfer of energy in the antennae via electron transport from chromophore to chromophore in a sequence of redox processes, as in the electron transport chains of photosynthesis or of mitochondrial respiration, could be excluded, since such an electron transport would need considerable activation energy. This is not the case, since a flux of excitation energy can be measured in the antennae at temperatures as low as 1 K. At these low temperatures light absorption and fluorescence still occur, whereas chemical processes catalyzed by enzymes are completely inactive. Under these conditions the energy transfer in the antennae pro-ceeds according to a mechanism that is related to those of light absorption and fluorescence.
When chromophores are positioned very close to each other, the quan-tum energy of an irradiated photon is transferred from one chromophore to the next. One quantum of light energy is named a photon, one quantum of excitation energy transferred from one molecule to the next is termed an exciton. A prerequisite for the transfer of excitons is a specific position-ing of the chromophores. This is arranged by proteins, and therefore the chromophores of the antennae always occur as protein complexes.
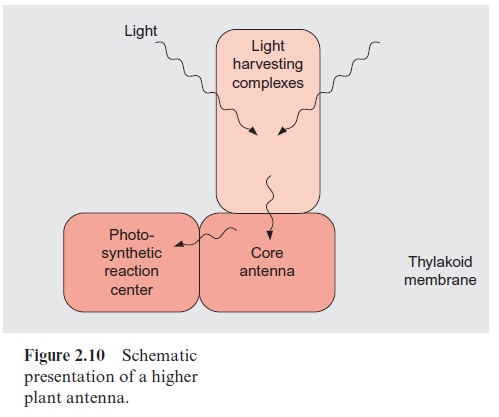
The antennae of plants consist of an inner part and an outer part (Fig. 2.10). The outer antenna, formed by the light harvesting complexes (LHCs), col-lects the light. The inner antenna, consisting of the core complexes, is an integral constituent of the reaction centers; it also collects light and con-ducts the excitons that were collected in the outer antenna to the photosyn-thetic reaction centers.
The LHCs are composed of polypeptides, which bind chl-a, chl-b, xan-thophylls, and carotenes. These proteins, termed LHC polypeptides, are encoded in the nucleus. A plant contains many different LHC polypeptides. In a tomato, for instance, at least 19 different genes for LHC polypeptides have been found, which are very similar to each other and are members of a multigene family. They are homologous, as they have all evolved from a common ancestor.
Plants contain two reaction centers, which are arranged in sequence: a reaction center of photosystem II (PS II), which has an absorption maxi-mum at 680 nm, and a photosystem I (PS I) with an absorption maximum at 700 nm. The function of these reaction centers will be described in sec-tions 3.6 and 3.8. Both photosystems are composed of different LHCs.
Related Topics