Chapter: Plant Biochemistry: Multiple signals regulate the growth and development of plant organs
Defense reactions are triggered by the interplay of several signals
Defense reactions are triggered by the interplay of several signals
Plants defend themselves against pathogenic bacteria and fungi by producing phytoalexins, and in some cases also by programmed cell death (hypersensitive reaction), in order to control an infection. Animals feeding on plants may stimulate the production of defense compounds, which make the plant poisonous or indigestible.
These various defense reactions are initiated by the interplay of several sig-nal components in a network. After an attack by pathogens or as a response to abiotic stress, signal cascades, including the phosphoinositol cascade (sec-tion 19.1), are induced, which lead to an increase of the Ca2+ concentration in the cytosol, which activates the Ca2+ -dependent protein kinases (CDK). This in turn activatesprotein kinase cascades, which modulate gene expression via transcription factors . Moreover, in an early response, super-oxide (•O2–) and/or H2O2 (reactive oxygen species (ROS)) are synthesized by an NADPH oxidase located in the plasma membrane. The ROS repre-sent chemical weapons for a direct attack on the pathogens, but they are also signal components for inducing signal cascades to initiate the production of other defense compounds. H2O2 is involved in the lignification process and thus plays a role in the solidification of the cell wall, another defense strategy against pathogens.
The formation of nitric oxide (•NO), a radical, is a further early response to pathogen attack. It is known as a signal component in animals and plants, and is released by the oxidation of arginine, which is catalyzed by nitric oxide synthase.
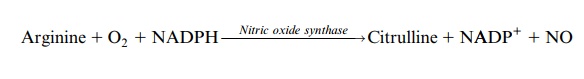
Alternatively, NO can be formed from nitrite in a side-reaction of the nitrate reductase.

NO is an important messenger in hormonal responses in plants and is involved in the defense against biotic and abiotic stress. NO is an impor-tant signal component that regulates, via cGMP and ADP ribose, Ca2+ channels to increase the cytoplasmic Ca2+concentration from intracellular stores, and in this way activates signal cascades. In connection with MAPK cascades, it promotes the synthesis of phytoalexins and is involved in the initiation of programmed cell death. In addition to abscisic acid, it induces the opening of stomata .
A precise control of the NO concentration in the cell is needed for it to function as a signal component. The regulation of its synthesis is to a large extent still unknown. A glutathione-dependent formaldehyde dehy-drogenase (FALDH) is involved in the control of the cellular NO concen-tration, by which NO is reversibly bound to glutathione. NO is eliminated by oxidation to nitrate.
Salicylic acid and jasmonic acid are signal molecules in pathogen defense
Jasmonic acid and salicylic acid are both involved in signal cascades induced during pathogen attack. SA plays a crucial role in defense responses against biotrophic pathogens (which keep the cell alive), and hemi-biotrophic pathogens (which initially keep the cell alive but kill them at a later stage). Mutants of transgenic tobacco plants, where the synthesis of salicylic acid had been intercepted, proved to be very vulnerable to infections by biotrophic and hemi-biotrophic pathogens. Enzymes induced by salicylic acid include β1.3 glucanase, which digests the cell wall of fungi, and lipoxygenase, a crucial enzyme in the pathway of the synthesis of jasmonic acid (Figs. 15.29 and 15.30). JA and ethylene are involved in the defense against herbivorous insect and necrotrophic patho-gens (which kill cells).
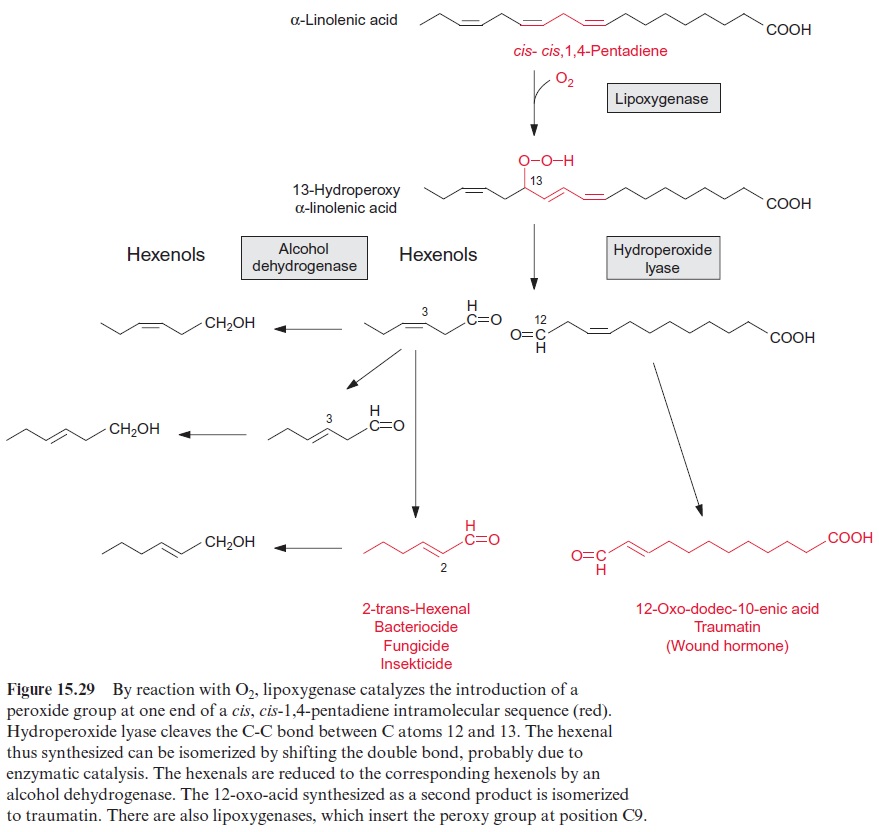
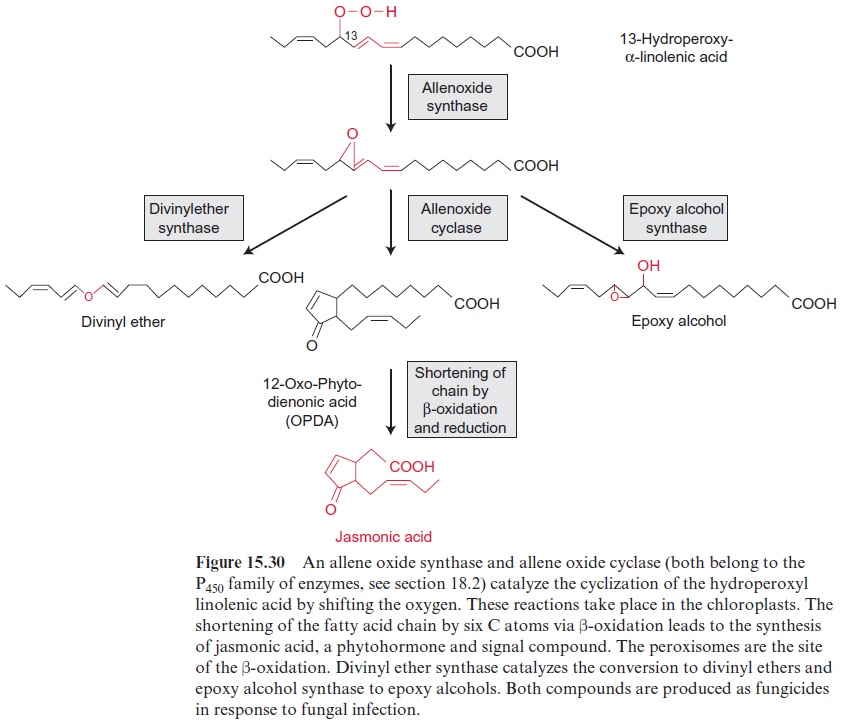
A number of components of the SA signaling cascade which regulate gene expression via transcription factors have by now been identified. Also for JA main signaling components are known, which interact with tran-scription factors. Both the SA and JA defense pathways contain different components, between which there is, however, positive and negative cross talk. Plant hormone signaling pathways are not isolated pathways but are interconnected with complex regulatory networks involving various defense signaling pathways and developmental processes. A better understanding of phytohormone-mediated plant defense responses is important in design-ing effective strategies for engineering crops for disease and pest resistance.
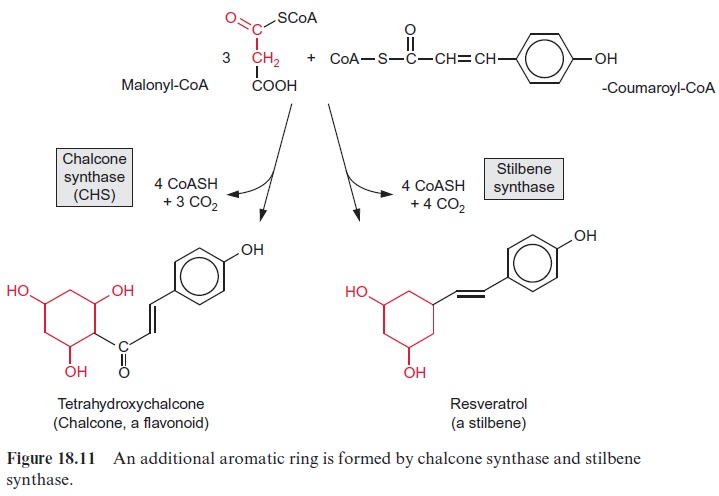
JAs are involved in diverse processes such as seed germination, root growth, tuber formation, tendril coiling, fruit ripening, leaf senescence and stomatal opening. Jasmonic acid regulates the development of pollen in some plants. Arabidopsis mutants, which are unable to synthesize jasmonic acid, cannot produce functioning pollen and therefore are male-sterile. The formation of jasmonic acid (e.g., induced by systemin) is regulated by a sig-nal cascade involving Ca2+ ions and MAP kinases. Also in the perception of jasmonic acid a MAP kinase cascade regulating transcription factors appears to be involved. Jasmonic acid, its methylester and its precursor 12-oxo-phytodienoic acid (OPDA) play a central role in defense reactions. As a response to fungal infection, jasmonic acid induces the synthesis of pheny-lammonium lyase (PAL) (Fig. 18.2), the entrance enzyme of phenyl propa-noid biosynthesis andchalcone synthase (CHS) (Fig.18.11), the key enzyme of flavonoid biosynthesis. As a response to wounding by herbivores, jasmonic acid causes plants to produce proteinase inhibitors. As a response to mechanical stress (e.g., by wind), jasmonic acid triggers the elevated growth in the thickness of stems or tendrils to make the plants more stable.
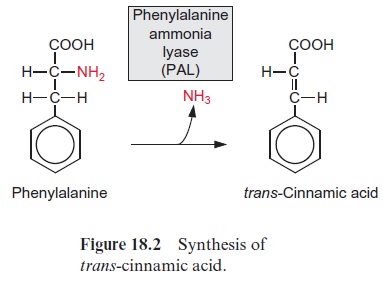
In many cases an attack by herbivores initiates defense responses not only in the wounded leaves, but also in more distant parts of a plant (systemic response). This requires a long-distance transport of signal sub-stances within the plant. Recent results indicate thatjasmonic acid and methyl jasmonate function as such a systemic wound signal in establishing a sys-temic acquired resistance (SAR). Also methyl salicylate is responsible for a long-distance signal transfer.
Related Topics