Chapter: Human Nervous System and Sensory Organs : The Ear
Vestibular Sensory Cells - Structure of The Ear
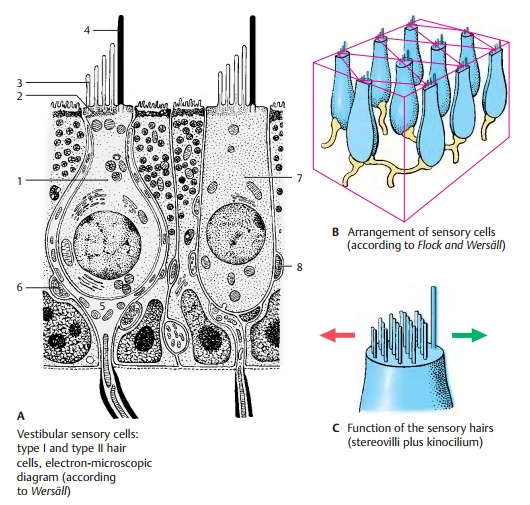
Vestibular Sensory Cells
The hair cells of the acoustic maculae and ampullary crests share the same structural principle. They are mechanoreceptors re-sponding to the tangential deflection of their sensory hairs. There are two types of vestibular hair cells: type I cells have the shape of a flask, while type II cells have the shape of a cylinder.
Type I hair cells (A1) have a round cell bodyand a narrow neck; below their apical sur-face lies a dense terminal web (A2). The api-cal cell surface is differentiated into ap-proximately 60 stereovilli (A3) of graduated lengths and a single, very long motile kinocilium(A4) with a basal body at itsorigin. Each type I hair cell is surrounded on its lateral and basal surfaces by a nerve calyx (A5), which is formed by a thick nerve fiber. The upper part of the calyx contains vesicles and closely adjoins the hair cell; this region is therefore regarded as the syn-aptic part of the hair cell. The nerve calyx, in turn, is contacted by heavily granulated nerve endings (A6), possibly representing the terminals of efferent nerve fibers.
Type II hair cells (A7) are equipped with anidentical set of sensory hairs. At the base of the cell lie more (A8) or less heavily granu-lated terminals of nerve fibers.
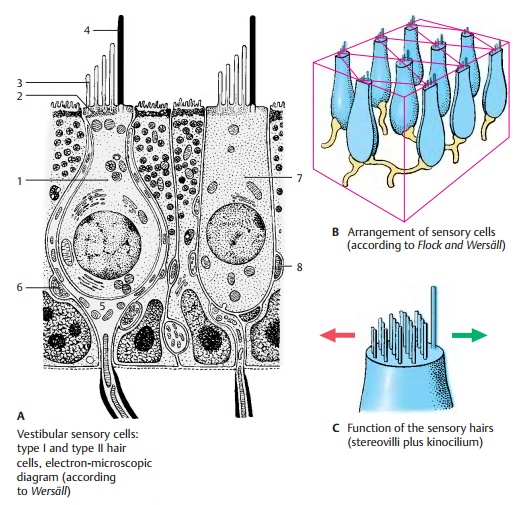
All hair cells of a sensory field show a uni-form orientation of their kinocilia (B). Elec-trophysiological studies have shown that bending of the stereovilli toward the kinocilium results in stimulation (green arrow), while bending in the opposite direc-tion leads to inhibition (red arrow) (C). Movements in intermediate directions cause stimulation or inhibition below the threshold of sensation.
In this way, the vestibular apparatus is able to register each movement precisely. The semicircular ducts (kinetic labyrinth) controlprimarily the eye movements, whereas the acoustic maculae (tonic labyrinth) directly af-fect muscular tone, in particular, the tension of extensor muscles and neck muscles.
Spiral Ganglion and Vestibular Ganglion(D)
The spiral ganglion (D9) consists of a chain of nerve cell clusters lying in the modiolus at the margin of the osseous spiral lamina. Together these clusters form a spiral. They contain bipolar neurons, the peripheral processes (dendrites) of which extend to the hair cells of the organ of Corti. Their central processes (axons) run as foraminous spiraltract (D10) to the axis of the modiolus,where they unite to form the radixcochlearis(root of the cochlear nerve) (D11).
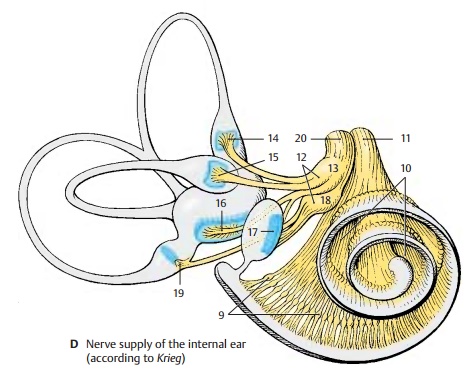
The vestibular ganglion (D12) lies at the floor of the internal acoustic meatus. It con-sists of a superior part and an inferior part. The bipolar neurons of the superior part (D13) send their peripheral processes to the ampullary crests of the anterior semicircu-lar duct (D14) (anterior ampullary nerve) and the lateral semicircular duct (D15) (lateral ampullary nerve), to the macula of the utricle (D16) (utricular nerve) and to part of the macula of the saccule (D17). The neurons of the inferior part (D18) supply the ampullary crest of the posterior semicircu-lar duct (D19) (posterior ampullary nerve) and part of the macula of the saccule (saccu-lar nerve). The central processes form the radix vestibularis(root of the vestibularnerve) (D20), which together with the radix cochlearis extends in a common nerve sheath through the internal acoustic meatus into the middle cranial fossa.
Related Topics