Chapter: Human Nervous System and Sensory Organs : The Ear
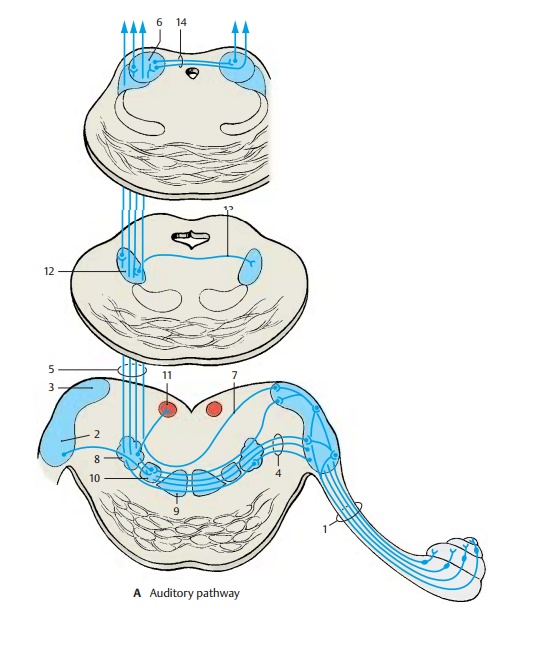
Auditory Pathway of The Ear
Auditory Pathway
Cochlear Nuclei (A, B)
The fibers of the radix cochlearis (A1) enter the medulla oblongata at the level of the anterior cochlear nucleus (AB2) and bifurcate.The ascending branches extend to the poste-rior cochlear nucleus (AB3), and the de-scending branches to the anterior cochlear nucleus. The projection of the cochlea onto the nuclear complex is highly organized; fibers from the basal cochlear convolutions terminate in the dorsomedial parts of the nuclei, while fibers from the uppermost convolutions terminate in the ventrolateral parts. This regular distribution of the affer-ent fibers is the basis for the subdivision of the cochlear nuclei according to tonal frequencies (see p. 370, frequency analysis in the cochlea).
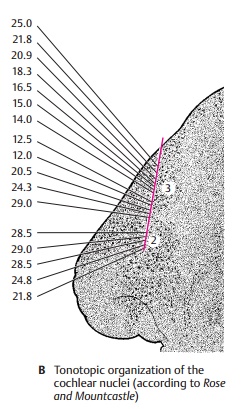
Such a tonotopic organization of the cochlear complex can be demonstrated by electrical recordings in an experimental an-imal (cat) (B). Recordings from individual neurons and simultaneous sonication with different sounds can detect the frequency at which an individual cell responds best. The frontal section through the oral region of the cochlear nuclei shows how the elec-trode, which scans from top to bottom, reg-isters in the posterior nucleus (B3) the recorded points in a precise sequence from high to low frequencies. Here lie the neu-rons for specific tonal frequencies in a regular sequence. When the electrode enters the anterior nucleus (B2), this sequence sud-denly stops and frequencies oscillate only in a specific range.
The secondary fibers of the auditory path-way originate from the neurons of the cochlear nuclei. Fiber bundles from the anterior cochlear nucleus cross to the op-posite side as a broad fiber plate mixed with nerve cells, the trapezoid body (A4), and then ascend as the lateral lem-niscus(A5) to the inferior col-liculus (A6). Fibers originating from the pos-terior cochlear nucleus cross obliquely as posterior acoustic striae(A7). A large portionof the lemniscus fibers run from the cochlear nuclei directly to the inferior col-liculus. A considerable number of fibers, however, are relayed to tertiary fibers in the intermediate nuclei of the auditory path-way, namely, in the posterior nucleus of thetrapezoid body (superior olive) (A8), in theanterior nucleus of the trapezoid body (A9), andin the nuclei of the lateral lemniscus. A tono-topic organization has been demonstrated in the nucleus of the posterior trapezoid body. The accessory nucleus lying medially to it, the medial nucleus of the superior olive (A10), receives fibers from the cochlear nu-clei of both sides and is interposed into a fiber system serving directional hearing. The fiber connections from the posterior nu-cleus of the trapezoid body to the abducens nucleus (A11) (reflex eye movements result-ing from the sensation of sound) are still dis-puted. The fibers are thought to extend be-yond the abducens nucleus to the con-tralateral cochlear nucleus and terminate as efferent fibers on the hair cells of the organ of Corti. They probably control the inflow of stimuli. The lateral nuclei of the lemniscus are scattered clusters of nerve cells inter-posed in the course of the lateral lemniscus. From the posterior nucleus of the lateral lem-niscus(A12), fibers cross to the con-tralaterallemniscus (Probst ’s commissure) (A13).
Inferior Colliculus (A)
The predominating portion of the lateral lemniscus terminates in the principal nu-cleus of the inferior colliculus in a topical pattern. Electrophysiological studies have demonstrated a tonotopic organization of this nucleus. It is a relay station for acoustic reflexes, from where acoustico-optic fibers extend to the superior colliculi and tec-tocerebellar fibers to the cerebellum. The inferior colliculi are interconnected by thecommissure of the inferior colliculi(A14).
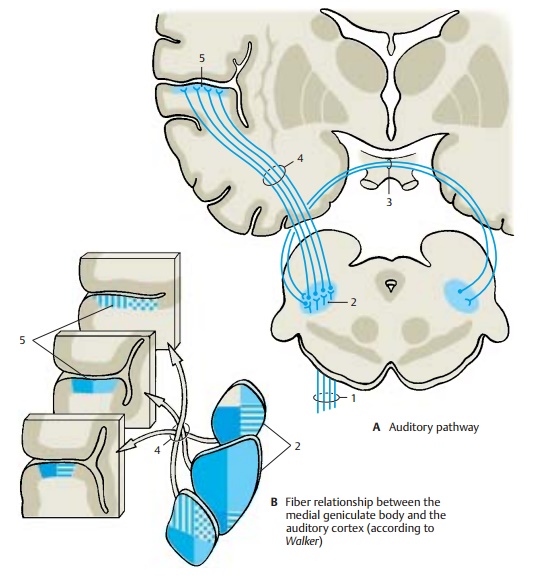
Medial Geniculate Body (A, B)
The next section of the auditory pathway is the peduncle of the inferior colliculi (A1), which extends as a strong fiber tract from the inferior colliculus to the medial geniculatebody (AB2), from where theacoustic radia-tionoriginates. The medial geniculate bodyis thought also to contain somatosensory fibers from the spinal cord as well as cere-bellar fibers. Obviously, it is not solely a relay station of the acoustic system but is also involved in other systems. Some fiber bundles of the peduncle of the inferior col-liculi are derived from the trapezoid nuclei and reach the medial geniculate body without synapsing in the inferior colliculus. The two medial geniculate bodies are thought to be interconnected by crossing fibers running through the inferior supra-optic commissure (Gudden’s commissure)(A3). It is uncertain whether such connec-tions exist in humans. However, the pres-ence of fibers descending from the auditory cortex and terminating in the geniculate body has been confirmed.
Acoustic Radiation (A, B)
The fibers of the acoustic radiation (AB4) run from the medial geniculate body trans-versely through the inferior posterior part of the internal capsule and ascend vertically in the temporal lobe to the auditory cortex. The fibers show a topical organization with individual sections of the medial geniculate body projecting to specific regions of the auditory cortex. Within the acoustic radia-tion, the fibers undergo a spiral rotation so that the rostral parts of the geniculate body project to the caudal cortical areas and the caudal parts of the geniculate body project to rostral cortical areas (B). This rotation has been demonstrated experimentally in monkeys and during maturation of the my-elin sheath in humans.
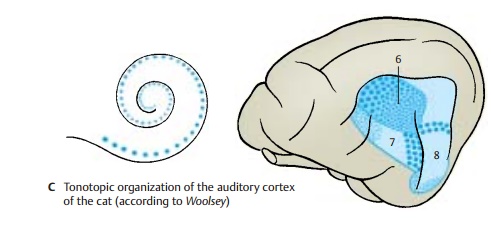
Auditory Cortex (A – C)
Electrical recordings from the cortex of various experimental animals (cat, monkeyunder simultaneous exposure to sounds of different frequencies have uncovered the tonotopic organization of the auditory cor-tex (AB5, C), where the uncoiled cochlea is represented from the basal convolution to the cupula. Three regions of hearing have been demonstrated: the primary auditoryarea (AI) (C6), the secondary auditory area (AII)(C7), and the area of the posterior ectosylviangyrus(Ep) (C8). In the primary auditory area,the neurons responding best to high frequencies (large blue dots) lie rostrally; the neurons responding best to low frequencies (small blue dots) lie caudally. In the secondary auditory area, the frequency sensitivity is arranged in the reverse order. Auditory area AI is the primary terminal ofthe acoustic radiation, while auditory areasAII and Ep are regarded assecondary audi-tory areas. The relationships may be com-pared to those of the visual cortex, where area 17 is the terminal of the optic radiation while areas 18 and 19 are secondary inte-gration areas. Auditory area AI corresponds in humans to area 41, which covers Heschl’s convolutions (transverse temporal gyri) and is the terminal of the acoustic radiation. Areas 42 and 22 are secondary auditory areas and include Wernicke’s speech center for the comprehension of lan-guages. Hence, the auditory cor-tex must be regarded as a region much larger than Heschl’s convolutions.
The auditory pathway has several commis-sural systems along its course that provide fiber exchanges at various levels. Some fiber bundles also ascend to the ipsilateralaudi-tory cortex. The latter therefore receives im-pulses from both organs of Corti, and this is especially important for directional hearing.
Related Topics