Chapter: Human Nervous System and Sensory Organs : The Ear
Vestibular Pathways of The Ear
Vestibular Pathways
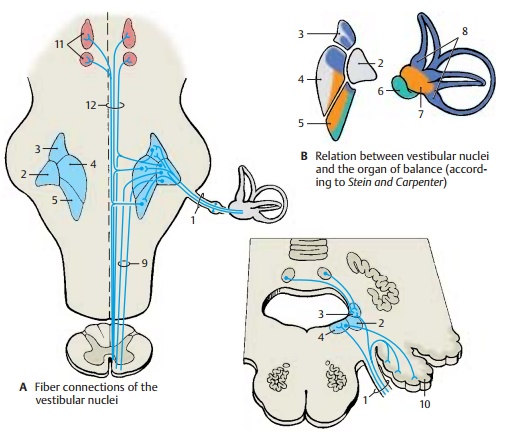
Vestibular Nuclei (A, B)
The fibers of the radix vestibularis (A1) enter the medulla oblongata at the level of the lateral vestibular nucleus (Deiters’ nucleus)(AB2) and bifurcate into ascending and de-scending branches that terminate in the su-perior vestibular nucleus (Bechterew’s nu-cleus) (AB3), in the medial vestibular nucleus(Schwalbe’s nucleus) (AB4), and in the infe-rior vestibular nucleus (AB5). Thenerve fibers for different parts of the laby-rinth extend to specific regions of the nu-clear complex. Fiber bundles for the maculaof the saccule(B6) terminate in the lateralpart of the inferior nucleus, while fibers for the macula of the utricle (B7) end in the me-dial part of the inferior nucleus and in the lateral part of the medial nucleus. Fibers for theampullary crests (B8) terminate pri-marily in the superior nucleus and in the upper part of the medial nucleus.
Certain groups of neurons respond to linear acceleration and others to rotational accel-eration. Some neurons respond to ipsilateral rotation, others to contralateral rotation. The vestibular complexes of both sides are interconnected by commissural fibers, through which some nerve cell clusters are stimulated by the labyrinth of the contralateral side. In addition to labyrinthine fibers, cerebellar fibers from the vermis and the fastigial nuclei as well as spi-nal fibers transmitting impulses from the joint receptors terminate in the nuclear complex. Efferent fibers for central control run from the vestibular nuclei back to the sensory epithelia.
Secondary Vestibular Pathways (A, C)
These are connections to the spinal cord, to the reticular formation, to the cerebellum, and to the oculomotor nuclei. The vesti-bulospinal tract (A9) originates from the neu-rons of Deiters’ nucleus (lateral vestibular nucleus) and reaches into the sacral spinal cord. Its fibers terminate at the spinal inter-neurons and activate the α- and γ-mo-toneurons of the extensor muscles.
The numerous fibers extending to the retic-ular formation stem from all vestibular nu-clei. To the cerebellum run direct fibers from the vestibular ganglion as well as fiber bundles from the medial and inferior vestibular nuclei. They terminate in the nodulus and flocculus (A10) and in parts of the uvula (vestibulocerebellum, p. 152, A6; p. 164, B). The fibers ascending to the oculo-motor nuclei (AC11) stem primarily from the medial and superior vestibular nuclei; they form part of the medial longitudinal fasciculus (A12). There is also a vesti-bulocortical connection through the thalamus (ventral intermediate nucleus, p. 184, B13). Electrophysiological studies re-sulted in a projection of vestibular impulses to a small region in the anterior postcentral area near the facial region of the sensory homunculus.
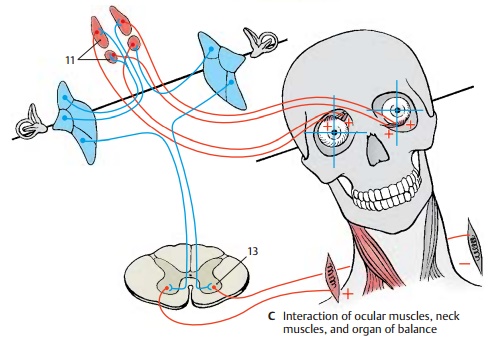
Interaction of eye muscles, neck muscles, and organ of balance (C). The connectionbetween vestibular complex and oculomo-tor nuclei brings circumscribed groups of neurons into contact with each other. Nerve cell clusters receiving impulses from a specific semicircular duct are probably con-nected with nerve cell clusters innervating a specific eye muscle. This would explain the exceptionally precise interaction of vestib-ular apparatus, eye muscles, and neck muscles, thus permitting fixation of an ob-ject even during movements of the head. We always perceive a stationary, vertical image of our surroundings despite our head movements. To guarantee such a constant visual impression, each head movement is compensated for by rotation of the eyeballs. The finely tuned interaction of neck muscles and ocular muscles is controlled by the ves-tibular apparatus via γ-motoneurons (C13).
Related Topics