Chapter: Medical Microbiology: An Introduction to Infectious Diseases: Bacterial Genetics
Transformation - Genetic Exchange in Bacteria
Transformation
Transformation was first demonstrated in 1928 by F. Griffith, a British public health officer, who showed that virulent, encapsulatedStreptococcus pneumoniae (pneumococci) that had been killed by heat could confer on living, avirulent, nonencapsulated pneumo-cocci the ability to make the polysaccharide capsule of the killed organisms and thus become virulent for mice. Subsequent work in 1944 by O. T. Avery, C. M. MacLeod, and M. McCarty at the Rockefeller Institute revealed that the “transforming factor” from the dead pneumococci was nothing other than DNA. This discovery had enormous impact on biology, because it was the first rigorous demonstration that DNA is the macromolecule in which genetic information is encoded. It opened the door to modern molecular genetics.
The ability to take up DNA from the environment is called competence, and in many species of bacteria, it is encoded by chromosomal genes that become active under certain environmental conditions. In such species, transformation can occur readily and is said to be natural. Other species cannot enter the competent state but can be made permeable to DNA by treatment with agents that damage the cell envelope making an artificial trans-formation possible.
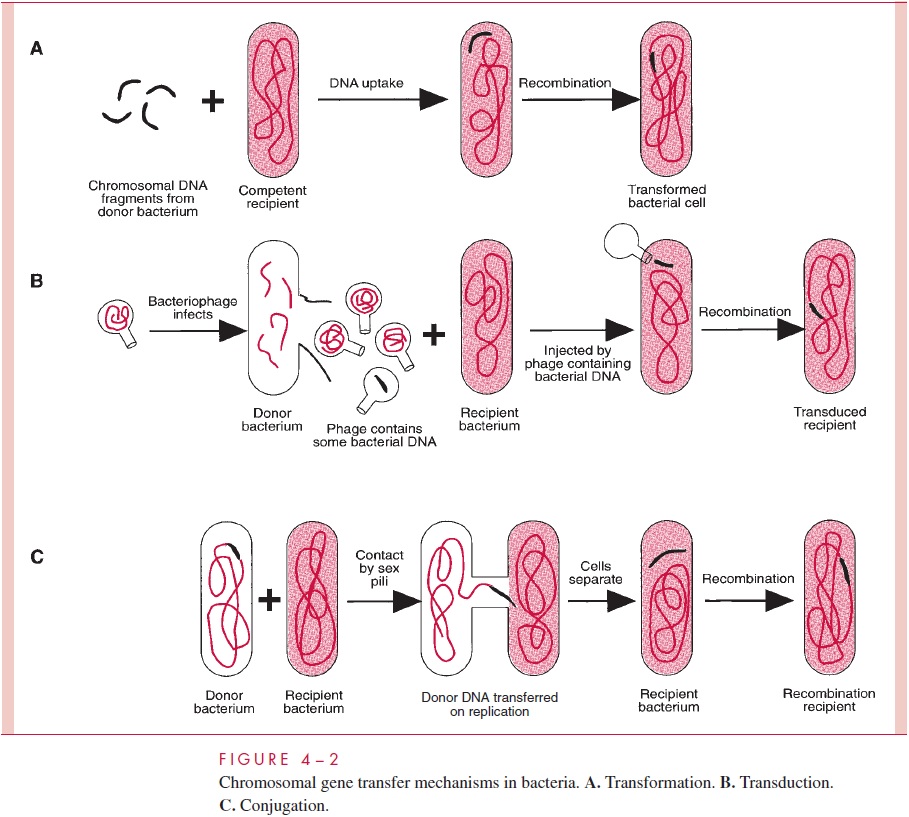
Natural transformation must be important in nature, judged by the variety of mecha-nisms that different bacteria have evolved to accomplish it. Two of the best-studied systems are those of the Gram-positive pneumococcus and a Gram-negative rod, Haemophilus in-fluenzae. Pneumococcal cells secrete a proteincompetence factorthat induces many of thecells of a culture to synthesize special proteins necessary for transformation, including an autolysin that exposes a cell membrane DNA-binding protein. Any DNA present in the medium is bound indiscriminately; even salmon sperm DNA can be bound and taken up as readily as DNA from another pneumococcal cell.
The surface-bound double-stranded DNA is cleaved into fragments of about 6 to 8 kilobases (kb). One strand is degraded by a nucle-ase, while the complementary strand of each fragment is taken up by a process that seems to be driven by the proton-motive force of the cell membrane . The fate of the internalized DNA fragment then depends on whether it shares homology (the same or simi-lar in base sequence) with a portion of the recipient cell’s DNA. If so, recombination can occur by a process described later, but heterologous DNA (no similarity to the endogenote) is degraded and causes no heritable change in the recipient.
Transformation in H. influenzae is somewhat different. There is no competence factor, and cells become competent merely by growth in an environment rich in nutrients. Only homologous DNA (ie, DNA from the same or a closely related species ofHaemophilus) is taken up, and it is taken up in double-stranded form. The selectivity is brought about by the presence of a special membrane protein that binds to an 11-base pair (bp) sequence (5'-AAGTGCGGTCA-3') that occurs frequently in Haemophilus DNA and infrequently in other DNAs. Following binding to molecules of this protein, the homologous DNA is internalized by a mechanism that resembles membrane invagination, resulting in the tem-porary residence of the exogenote in cytosolic membrane vesicles. Although the DNA taken up is double stranded, only one of the two strands participates in the subsequent re-combination with the endogenote.
The common use of E. coli as a host cell in which to clone genes on hybrid plasmids (see Invertible DNA Segments and Recombinational Regulation of Gene Expression) depends on procedures involving treatment with salt and temperature shocks to bring about artificial transformation; this organism has no natural competence mechanism. In contrast, the pathogen, Neisseria gonorrhoeae regularly uses transformation to bring about changes in the antigenic nature of its pili, as described later in the section on recombination.
Related Topics