Chapter: Medical Microbiology: An Introduction to Infectious Diseases: Bacterial Genetics
Transduction - Genetic Exchange in Bacteria
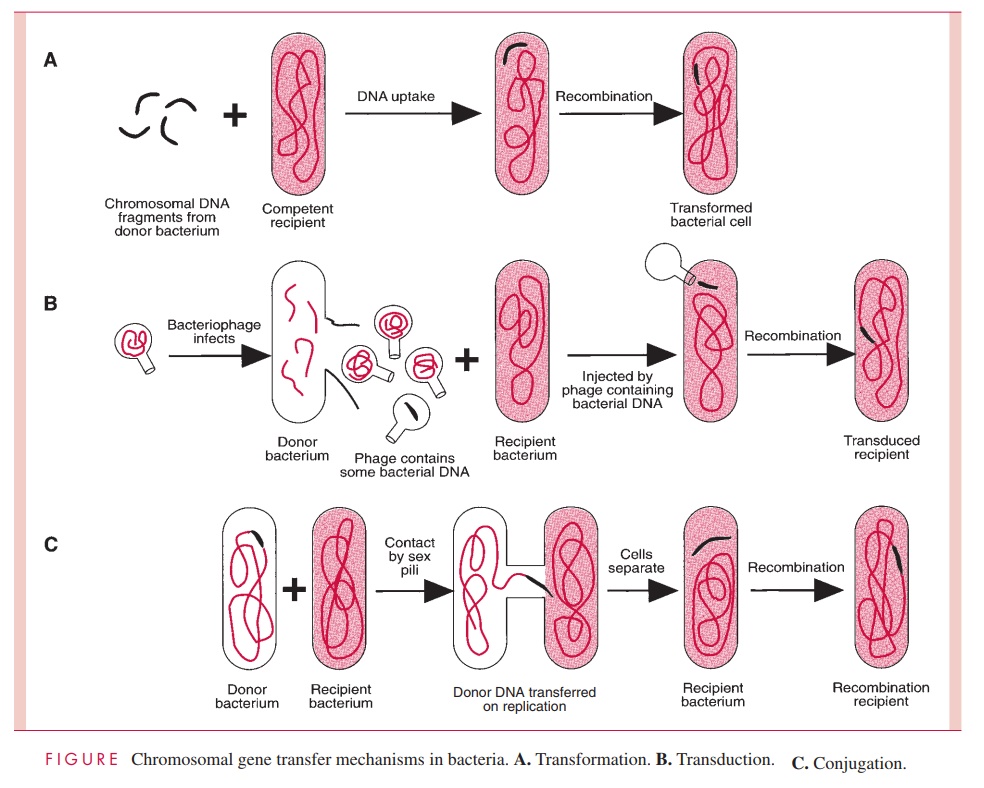
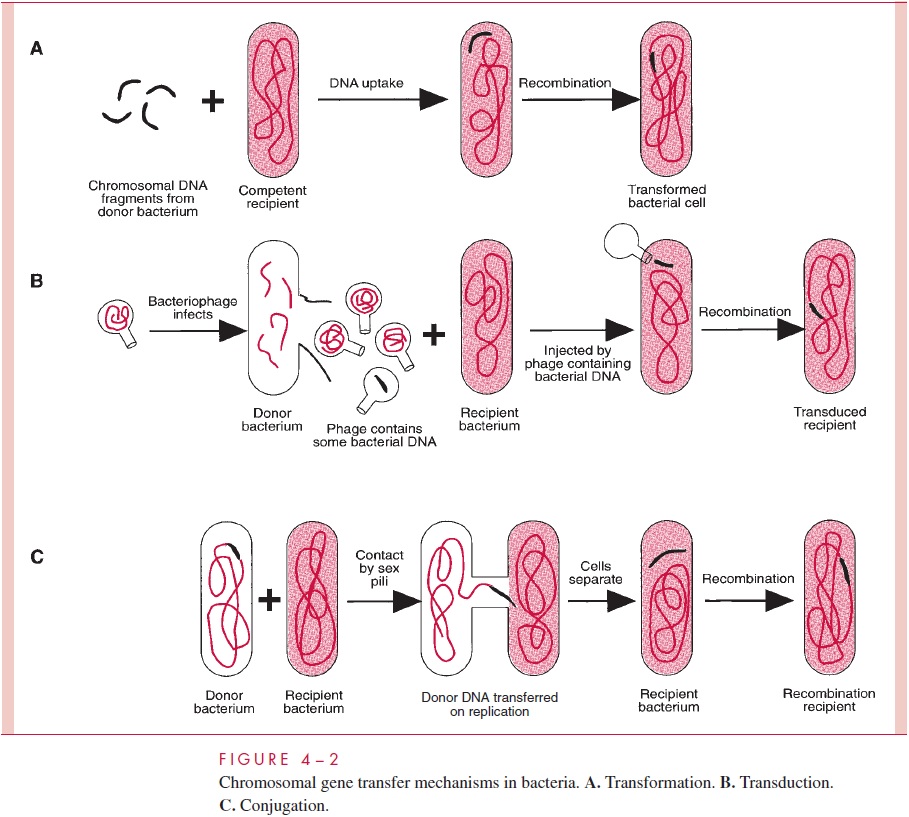
Transduction
Transduction is virus-mediated transfer of genetic information from donor to recipient cell. To understand transduction and its several mechanisms, it is necessary to preview the nature of bacterial viruses.
Viruses are capable of reproduction only inside living cells. Those that grow in bacte-ria are called bacteriophages, or simply phages. They are minimally composed of pro-tein and nucleic acid, although some may have a very complex structure and composition. The individual virus particle or virion consists of a protein capsid enclosing genomic nu-cleic acid, which is either RNA or DNA, but never both. Virions infect sensitive cells by adsorbing to specific receptors on the cell surface and then, in the case of phages, inject-ing their DNA or RNA. Phages come in two functional varieties according to what hap-pens after injection of the viral nucleic acid. Virulent (lytic) phages cause lysis of the host bacterium as a culmination of the synthesis of many new virions within the infected cell. Temperate phages may initiate a lytic growth process of this sort or can enter a qui-escent form (called a prophage), in which the infected host cell is permitted to proceed about its business of growth and division but passes on to its descendants a prophage genome capable of being induced to produce phage in a process nearly identical to the growth of lytic phages. The bacterial cell that harbors a latent prophage is said to be a lysogen (capable of producing lytic phages), and its condition is referred to as lysogeny.Lysogens are immune to infection by virions of the type they harbor as prophage. Occa-sionally, lysogens are spontaneously induced and lysed by the phage and release mature virions (as many as 75 to 150 or more per cell) into the environment. When triggered by UV irradiation or certain chemicals, an entire population of lysogens are induced simulta-neously to initiate reproduction of their latent virus followed by lysis of the host cells. In-fection of a sensitive cell with the temperate phage can lead to either lysis or lysogeny.
The prophage of different temperate phages exists in one of two different states. In the first, the prophage DNA is physically integrated into a bacterial chromosome; in the sec-ond, it remains separate from the chromosome as an independently replicating, circular-ized, molecule of DNA. Prophages of this sort are in fact plasmids.
For the most part, transduction is mediated by temperate phage, and the two broad types of transduction result from the different physical forms of prophage and the differ-ent means by which the transducing virion is formed. These are termed generalizedtransduction, by which any bacterial gene stands an equal chance of being transduced toa recipient cell, and specialized or restricted transduction, by which only a few genes can be transduced.
Generalized Transduction
Some phages package DNA into their capsids in a nonspecific way, the headful mecha-nism, in which any DNA can be stuffed into the capsid head until it is full. (The head is the principal structure of the virion to which, in some cases, a tail is attached;) An endonuclease then trims off any projecting excess. If fragments of host cell DNA are around during the assembly of mature virions, they can become packaged in place of virus DNA, resulting in pseudovirions. Pseudovirions are the transducing agents. They can ad-sorb to sensitive cells and inject the DNA they contain as though it were viral DNA. The result is the introduction of donor DNA into the recipient cell.
Any given gene has an equal probability of being transduced by this process. With the temperate phage P1 of E. coli, this probability is approximately one transduction event per 105 to 108 virions, because nearly 1 out of every 1000 phage particles made in a P1 lytic infection are pseudovirions, and the bacterial DNA fragments packaged are 1 to 2% of the length of the chromosome. Cotransduction of two bacterial genes by a single pseudovirion occurs only if they are located close together within this small length of the chromosome, and this fact facilitates mapping the position of a newly discovered gene.
Once injected into the host cell, the transduced DNA is lost by degradation unless it can recombine with the chromosome of the recipient cell, usually by homologous recom-bination (see below, Invertible DNA Segments and Recombinational Regulation of Gene Expression) in which both strands of the exogenote cross into and replace the homolo-gous segment of the recipient’s chromosome. However, sometimes the exogenote can per-sist without degradation by assuming a stable circular configuration.
Specialized Transduction
It has been noted that the prophage of some phages is integrated into the lysogen’s chro-mosome. This integration does not occur haphazardly but is restricted to usually one site, called the att (attachment) site. When a lysogen carrying such a prophage is induced to produce virions, excision of the viral genome from the bacterial chromosome occasion-ally (eg, in 1 of 105 to 106 lysogens) occurs imprecisely, resulting in a pickup of genes of the bacterium adjacent to the att site. The resulting virion may be infectious (if no essen-tial phage genes are missing) or defective (if one or more essential genes are missing). In either case, adsorption to a sensitive cell and injection of the DNA can occur, and integra-tion of the aberrant phage genome into the chromosome of the new host cell results in the formation of a lysogen containing a few genes that have been transduced as hitchhikers with the phage genome. Integration of the phage genome automatically accomplishes the recombinational event needed to guarantee reproduction of the transduced genes. Only genes that border the att site stand a chance of being transduced by this process, which is why it is called specialized or restricted transduction.
Because the original pickup event is rare, the first transducing process is termed low-frequency transduction; however, when a lysogenic transductant is, in turn, induced toproduce phage, all of the new virions carry the originally transduced bacterial gene. The resulting mixture of lysed cells and virions now brings about high-frequency transduc-tion of the attached genes.
Bacterial geneticists have learned to move genes of interest near the phage integration site and thereby construct specialized transducing phages containing these genes. Such transducing phages are valuable aids to cloning and sequencing genes and to studying their function and regulation. Obviously a temperate phage that could form a prophage by integrating randomly at any site in the bacterial chromosome would be of special use. The temperate phage Mu of E. coli has this property.
Although both generalized transduction and specialized transduction can be regarded as the result of errors in phage production, transfer of genes between bacterial cells by phage is a reasonably common phenomenon. It occurs at significant frequency in nature; for example, genes conferring antimicrobic resistance in staphylococci are often trans-duced from strain to strain in this way. The toxins responsible for the severe clinical symptoms of diphtheria and of cholera are encoded by genes transduced into Corynebac-terium diphtheriae and Vibrio cholerae, respectively. Transduction is also used exten-sively as a tool in molecular biology research.
Related Topics