Chapter: Human Nervous System and Sensory Organs : The Eye
Photoreceptors - Structure of the Eye
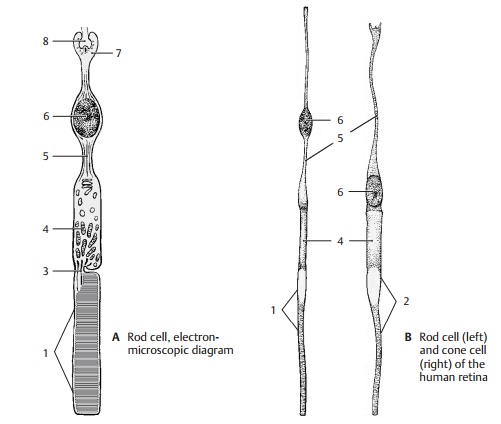
Photoreceptors
The
light-sensitive sensory cells have the same structural design in all
vertebrates. Next to the pigmented epithelium lies the outer segment of the photoreceptor cell; it ispartly buried in a
pigmented epithelial cell. The outer segment of rod cells is a cylinder (ABC1) containing several hundreds of
stacked, disk-shaped membrane pouches of uniform size. The outer segment of
cone cells (B2, D) is of a conical shape, and the proximal membrane folds are
larger than the distal ones. A thin, eccentric cyto-plasmic bridge, the connecting cilium (ACD3), links the outer segment to the innersegment (AB4). The
bridge contains a mod-ified cilium with 9 pairs of microtubules but without the
central pair characteristic of other cilia (see p. 285, D5, D6). The
connect-ing cilium is relatively long in some species, leaving a distinct space
between the two segments (A). In
humans, however, it is so short that both segments touch each other without
leaving a visible space between them (B).
The inner segment contains numerous Golgi stacks, ribosomes, and longitudinally
arranged mitochondria. The cell body then tapers to form an axonlike process (AB5) containing neurofilaments and
microtubules. The cell nucleus (AB6)
lies either at the transition from the inner segment to the axon or within the
axon. The cell terminates with an end bulb, the synap-tic terminal (A7).
In addition to the usualsynapses, the terminal develops invaginatedsynapses (A8)
in which the presynapticmembrane becomes invaginated and sur-rounds the
postsynaptic complex on all sides.
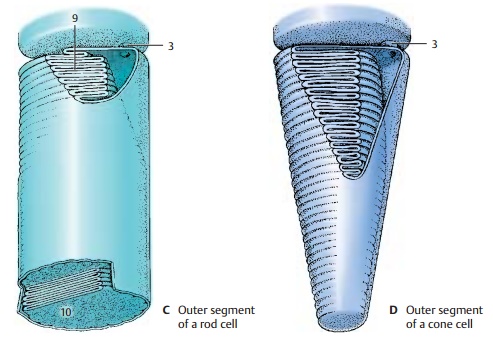
The
outer segment is the actual receptorpart of
the cell where the light is absorbed.The stacked membranes are formed by
in-foldings of the plasma membrane (C9)
in the proximal part of the outer segment. In the rod cells, they detach from
the outer membrane and form isolated disks in the distal part of the segment (C10). The visualpigment of the rod cells,rhodopsin,
is boundto the membrane of the disks. The forma-tion of visual pigment in the
inner segmentand its migration through the connecting bridge into the outer
segment can be fol-lowed using autoradiography by labeling the protein
component of rhodopsin with a radioactive amino acid. Once the labeled
substance has passed the bridge, it forms a band that migrates to the outer end
and then disappears (in rats within 10 days). The migrating band represents a
membrane disk that has incorporated labeled rhodop-sin. Thus, new disks are
being continuously formed in the rod cells, migrate to the distal end, and are
shed there. Fragments of the shed disks have been found in the pig-mented
epithelial cells. There is no new for-mation of disks in the outer segments of
cone cells (D). The infoldings of
the mem-brane are permanent and, in contrast to rod cells, there is no
detachment of membrane invaginations from the plasma membrane.
Only rod
cells contain rhodopsin. The ab-sorption of light changes the molecular
structure of rhodopsin, causing it to break down into its protein and pigment
com-ponents. From these components, rhodop-sin is continuously resynthesized in
the rods (rhodopsin–retinin cycle).
It absorbs light of all wavelengths and, thus, is not in-volved in color
vision. Rod cells are light–dark
receptors. The three different types ofcone cells each contain a different
pigment that absorbs only light of a specific wavelength. Cone cells are color receptors.
There
are some animal species in which the retina contains only cones, while it
contains only rods in other species (cat, cattle). Ani-mals with a retina
containing only rods can-not distinguish between colors. The bull, which is
said to react to red, is actually color blind.
Related Topics