Chapter: Human Nervous System and Sensory Organs : Basic Elements of the Nervous System
Neurotransmitters - The Synapse
Neurotransmitters
Transmission
of impulses at the chemical synapses is mediated by neurotransmitters. The most
widely distributed transmitter substances in the nervous system are acetylcholine (ACh), glutamate, gamma-aminobutyric acid (GABA), and glycine.Glutamate is the most common excitatory transmitter, GABA is
a transmitter of the in-hibitory synapses in the brain, and glycine is an
inhibitory transmitter in the spinal cord.
The catecholaminesnorepinephrine (NE) and dopamine
(DA) also act as transmitters, and so does serotonin
(5-HT). Many neu-ropeptides act not
only as hormones in thebloodstream but also as transmitters in the synapses
(e.g., neurotensin, cholecys-tokinin, somatostatin).
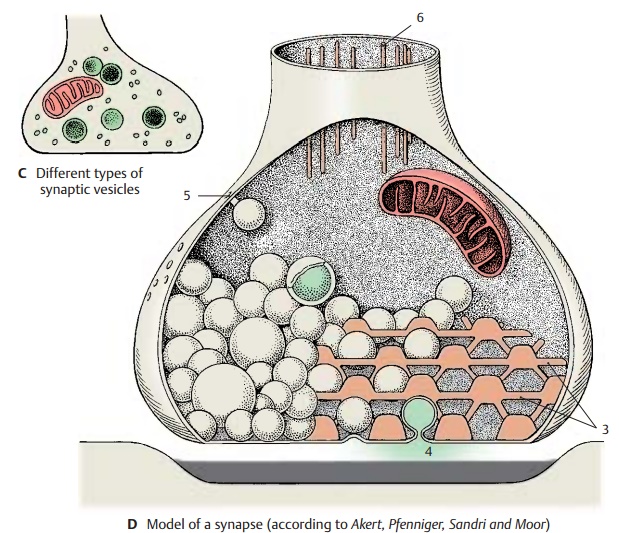
The
transmitters are produced in the peri-karyon and stored in the vesicles of the
axon terminals. Often only the enzymes required for transmitter synthesis are
produced in the perikaryon, while the transmitter sub-stances themselves are
synthesized in the boutons. The small and
clear vesicles are thought to carry glutamate and ACh, the elongated vesicles of the inhibitory synapsescarry
GABA, while norepinephrine and dopamine are present in the granular ves-icles (C).
Most
vesicles are located near the presyn-aptic membrane, the density of which can
be demonstrated by special procedures as a grid with hexagonal spaces (D3). The ves-icles pass through these
spaces to reach the presynaptic membrane and, upon excita-tion, empty their
content into the synaptic cleft by fusing with the presynaptic mem-brane (omega figure) (D4). The transmitter substances are delivered in certain quanta,
the morphological equivalents of which are the vesicles. Some of the
transmitter molecules return into the bouton by reup-take (D5).
D6
Axonal filaments.
Many
neurons, perhaps most of them, pro-duce more than one transmitter substance.
Nevertheless, they are classified according to their functionally most
important neu-rotransmitter as glutamatergic,
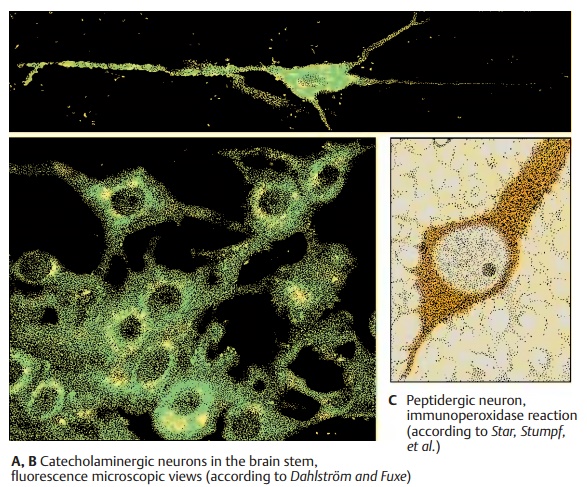
cholinergic, catecholaminergic (noradrenergic anddopaminergic), serotoninergic, and pepti-dergic neurons. The catecholaminergic and serotoninergic neurons can be
identifiedby fluorescence microscopy because their transmitters show a
green-yellow fluores-cence following exposure to formalin vapor (A, B). It is thus possible to trace the
axon and to recognize the perikaryon and the outline of its nonfluorescent
nucleus. The fluorescence is very faint in the axon, more distinct in the
perikaryon, and most intense in the axon terminals. It is here that the highest
concentration of transmitters oc-curs. Cholinergic
neurons can be demon-strated by a histochemical assay for acetyl-cholinesterase, the enzyme
required for thedegradation of acetylcholine. Since this enzyme is also
produced by noncholinergic neurons, the proper assay is by immunocy-tochemistry
using antibodies against cholineacetyltransferase,
the acetylcholine-synthe-sizing enzyme. Other transmitters and neu-ropeptides
can also be demonstrated by im-munocytochemistry (C). It has been shown by double-labeling that many neuropep-tides
are produced together with classical neurotransmitters within the same neuron.
So far, the functional significance of cotrans-mission,
i.e., the release of different trans-mitters by the same neuron, has been
stud-ied in detail only on some neurons of the au-tonomic nervous system.
Related Topics