Chapter: Essentials of Anatomy and Physiology: Muscular System
Muscle Contraction
Muscle Contraction
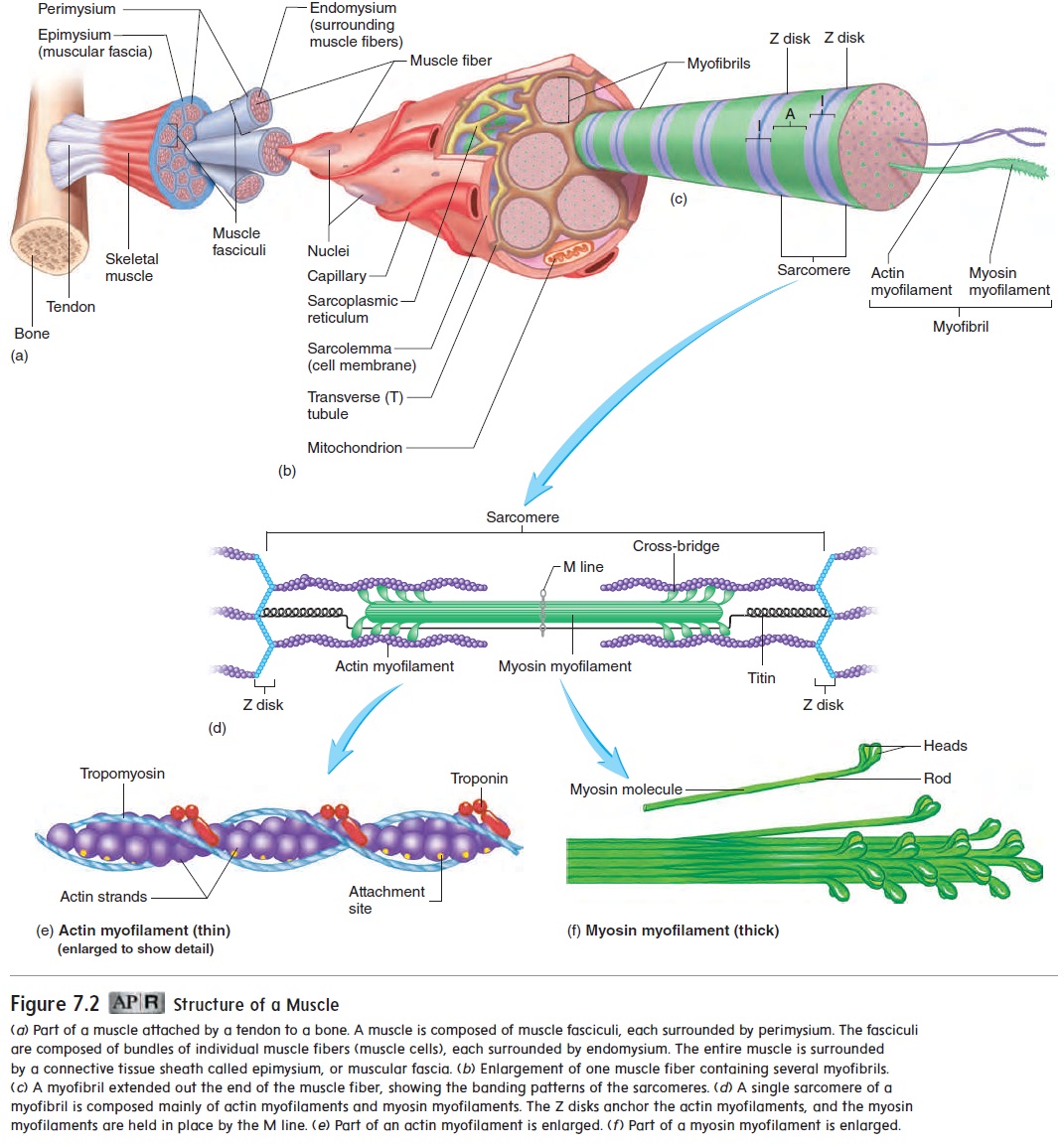
Contraction of skeletal muscle tissue occurs as actin and myosin myofilaments slide past one another, causing the sarcomeres to shorten. Many sarcomeres are joined end-to-end to form myofibrils. Shortening of the sarcomeres causes myofibrils to shorten, thereby causing the entire muscle to shorten.
The sliding of actin myofilaments past myosin myofilaments during contraction is called the sliding filament model of muscle contraction. During contraction, neither the actin nor the myosin fibers shorten. The H zones and I bands shorten during contraction, but the A bands do not change in length (figure 7.7).
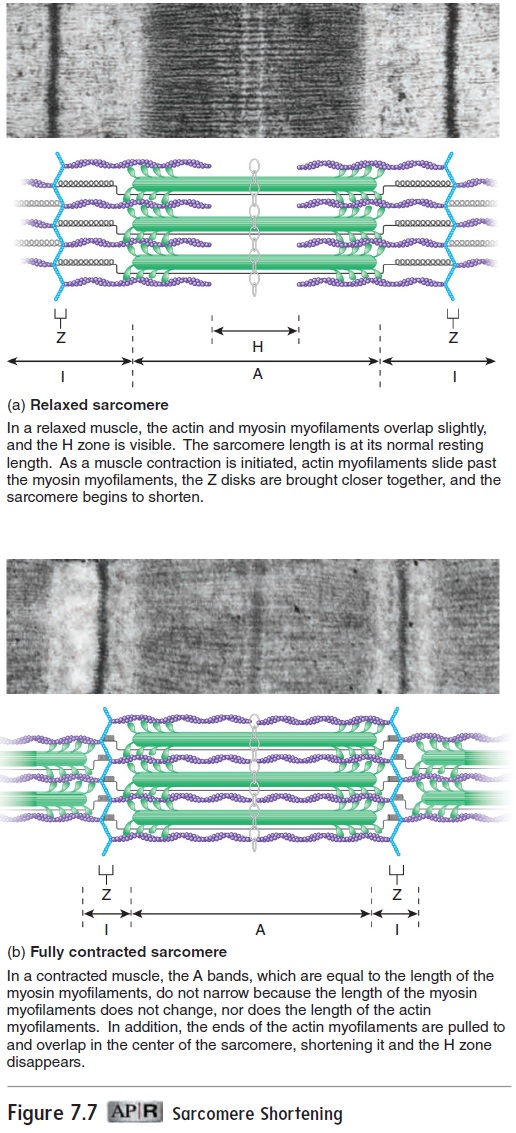
During muscle relaxation, sarcomeres lengthen. This length-ening requires an opposing force, such as that produced by other muscles or by gravity.
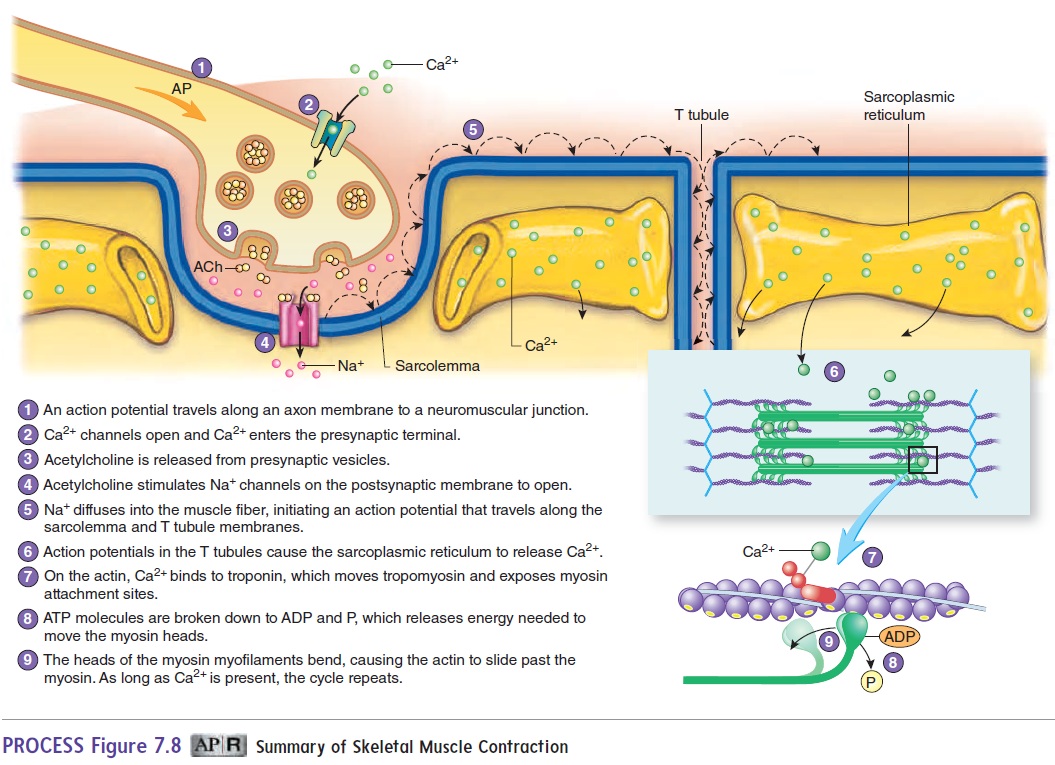
Figure 7.8 summarizes the sequence of events that occur as a stimulus causes a skeletal muscle to contract. Action potentials produced in skeletal muscle fibers at the neuromuscular junc-tion travel along the sarcolemma and the T tubule membranes (see figure 7.6). The action potentials cause the membranes of the sarcoplasmic reticulum adjacent to the T tubules to become more permeable to Ca2+, and Ca2+ diffuses into the sarcoplasm. The Ca2+ binds to troponin molecules attached to the actin myofilaments This binding causes tropomyosin molecules to move into a groove along the actin molecule, exposing myosin attachment sites on the actin myofilament. The exposed attachment sites on the actin myofilament bind to the heads of the myosin myofilaments to form cross-bridges between the actin and myosin myofilaments.
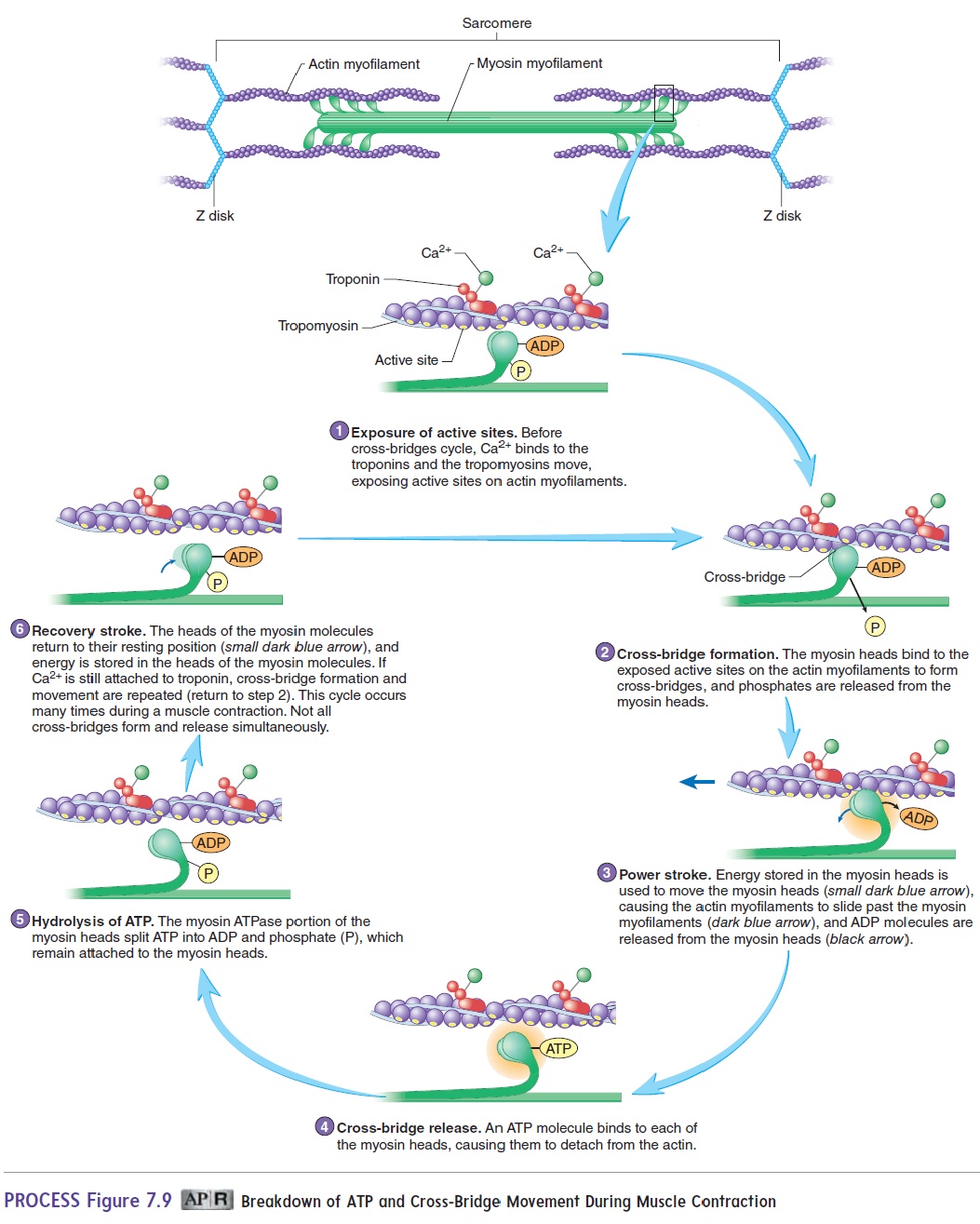
Energy for muscle contraction is supplied to the muscles in the form of adenosine triphosphate (ATP), a high-energy molecule produced from the energy that is released during the metabolism of food. The energy is released as ATP breaks down to adenosine diphosphate (ADP) and phosphate (P). During muscle contraction, the energy released from ATP is briefly stored in the myosin head. This energy is used to move the heads of the myosin myofilaments toward the center of the sarcomere, causing the actin myofilaments to slide past the myosin myofilaments. In the process, ADP and P are released from the myosin heads.
As a new ATP molecule attaches to the head of the myosin molecule, the cross-bridge is released, the ATP breaks down to ADP and P (which both remain bound to the myosin head), and the myosin head returns to its original position, where it can attach to the next site. As long as Ca2+ remains attached to tro-ponin, and as long as ATP remains available, the cycle of cross-bridge formation, movement, and release repeats (figure 7.9). A new ATP must bind to myosin before the cross-bridge can be released. After a person dies, ATP is not available, and the cross-bridges that have formed are not released, causing the muscles to become rigid. This condition is called rigor mortis (rig′er m̄o r′tis; stiffness + death).
Part of the energy from ATP involved in muscle contraction is required for the formation and movement of the cross-bridges, and part is released as heat. The heat released during muscle contraction increases body temperature, which explains why a person becomes warmer during exercise. Shivering, a type of generalized muscle contraction, is one of the body’s mechanisms for dealing with cold. The muscle movement involved in shivering produces heat, which raises the body temperature.
Muscle relaxation occurs as Ca2+ is actively transported back into the sarcoplasmic reticulum (a process that requires ATP). As a consequence, the attachment sites on the actin molecules are once again covered by tropomyosin so that cross-bridges cannot reform.
Muscle Twitch, Summation, Tetanus,and Recruitment
A muscle twitch is the contraction of a muscle fiber in response to a stimulus. Because most muscle fibers are grouped into motor units, a muscle twitch usually involves all the muscle fibers in a motor unit. A muscle twitch has three phases (figure 7.10). The lag phase, orlatent phase,is the time between the application of astimulus and the beginning of contraction. The contraction phase is the time during which the muscle contracts, and the relaxationphase is the time during which the muscle relaxes.
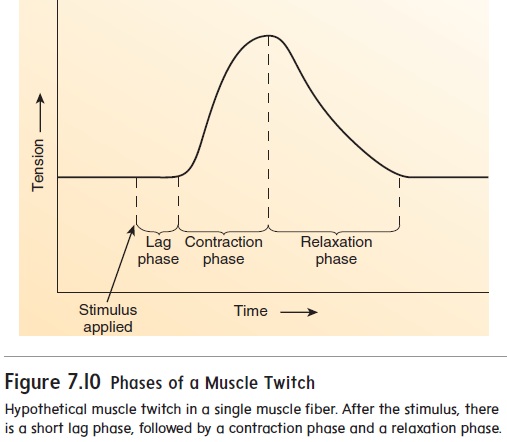
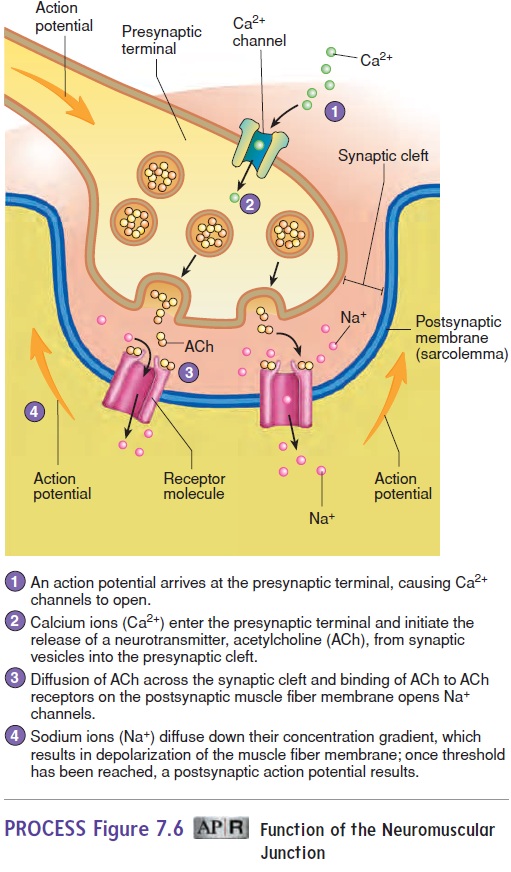
During the lag phase, action potentials are produced in one or more motor neurons. An action potential travels along the axon of a motor neuron to a neuromuscular junction (see figure 7.5). Once the stimulus reaches the neuromuscular junction, acetyl-choline must be released from the presynaptic terminal, diffuse across the synaptic cleft, and bind to receptors that allow the entry of Na+, which initiates an action potential on the postsynaptic membrane (see figure 7.6). Before the contraction phase can occur, the action potential must result in the release of Ca2+ from the sarcoplasmic reticulum and the formation of cross-bridges (see figure 7.9, steps 1–2).
The contraction phase results from cross-bridge movement and cycling (see figure 7.9, steps 3–6), which increases the tension produced by the muscle fibers (see figure 7.10).
During the relaxation phase, Ca2+ is actively transported back into the sarcoplasmic reticulum. As Ca2+ diffuses away from the troponin molecules, tropomyosin molecules once again block the attachment sites. Cross-bridge formation is prevented, and the ten-sion produced by the muscle fibers decreases (see figure 7.10).
The strength of muscle contractions varies from weak to strong. For example, the force muscles generate to lift a feather is much less than the force required to lift a 25-pound weight. The force of contraction a muscle produces is increased in two ways:(1) Summation involves increasing the force of contraction of the muscle fibers within the muscle, and (2) recruitment involves increasing the number of muscle fibers contracting.
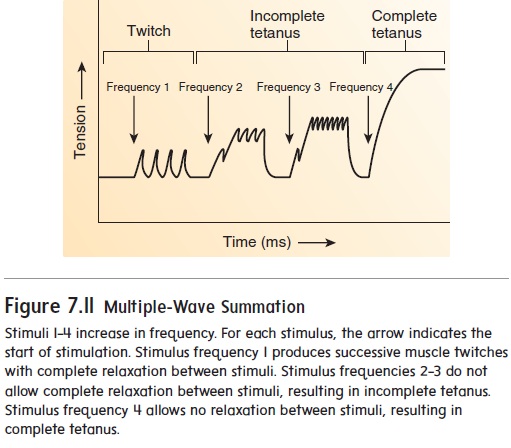
In summation, the force of contraction of individual muscle fibers is increased by rapidly stimulating them. When stimulus fre-quency, which is the number of times a motor neuron is stimulated per second, is low, there is time for complete relaxation of muscle fibers between muscle twitches (figure 7.11, stimulus frequency 1). As stimulus frequency increases (figure 7.11, stimulus frequencies 2–3), there is not enough time between contractions for muscle fibers to relax completely. Thus, one contraction summates, or is added onto, a previous contraction. As a result, the overall force of contraction increases.Tetanus (tet′a-nus; convulsive tension) is a sustained contraction that occurs when the frequency of stimula-tion is so rapid that no relaxation occurs (figure 7.11, stimulus frequency 4). It should be noted, however, that complete tetanus is rarely achieved under normal circumstances and is more commonly an experimentally induced muscular response. The increased force of contraction produced in summation and tetanus occurs because Ca2+ builds up in myofibrils, which promotes cross-bridge forma-tion and cycling. The buildup of Ca2+ occurs because the rapid production of action potentials in muscle fibers causes Ca2+ to be released from the sarcoplasmic reticulum faster than it is actively transported back into the sarcoplasmic reticulum.
In recruitment, the number of muscle fibers contracting is increased by increasing the number of motor units stimulated, and the muscle contracts with more force. When only a few motor units are stimulated, a small force of contraction is produced because only a small number of muscle fibers are contracting. As the number of motor units stimulated increases, more muscle fibers are stimulated to contract, and the force of contraction increases. Maximum force of contraction is produced in a given muscle when all the motor units of that muscle are stimulated (recruited).
If all the motor units in a muscle could be stimulated simulta-neously, the resulting motion would be quick and jerky. However, because the motor units are recruited gradually, some are stimulated and held in tetanus while additional motor units are recruited; thus, contractions are slow, smooth, and sustained. In the same way, smooth relaxation of muscle occurs because some motor units are held in tetanus while other motor units relax.
Fiber Types
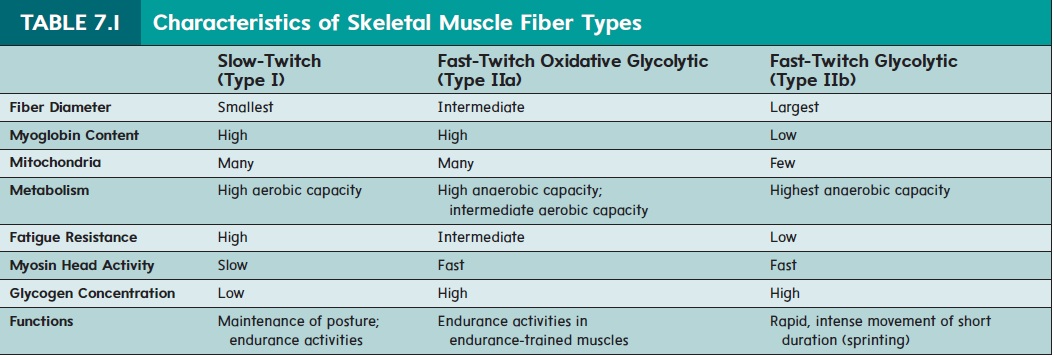
Muscle fibers are sometimes classified as either slow-twitch or fast-twitch fibers (table 7.1). This classification is based on differ-ences in the rod portion of the myosin myofilament (see figure 7.2). Slow-twitch fibers contain type I myosin as the predominant or even exclusive type. Fast-twitch fibers contain either type IIa or type IIb myosin myofilaments. Each of these three myosin types is the product of a different myosin gene.
The fast and slow names of these fibers indicate their contrac-tion speeds. Fast-twitch muscle fibers contract quickly, whereas slow-twitch muscle fibers contract more slowly. Among the fast-twitch fibers, type IIb fibers are the fastest and type IIa fibers contract at an intermediate speed. The type IIb fibers can contract ten times faster than slow-twitch (type I) fibers. However, while slower, the slow-twitch fibers can sustain the contraction for longer than the fast-twitch fibers. Likewise, type IIa fibers can sustain contractions longer than type IIb, but not as long as type I
Related Topics