Chapter: Introduction to Human Nutrition: Nutrition and Metabolism of Lipids
Lipoprotein structure: a shopping bag and groceries
Circulating lipids: lipoprotein structures and metabolism
Circulating blood lipids are insoluble in water and must be solubilized for transportation in the extracel-lular fluid by combining with bipolar molecules with charged and uncharged regions (apoproteins and phospholipids). This property, known as amphipath-icity, enables these molecules to associate with aqueous (hydrophilic) and nonaqueous (hydrophobic) envi-ronments and thus renders them perfect for enveloping insoluble lipids, chiefly TAG and CE, in macromolecular lipid–protein complexes called lipo-proteins. It is worth remembering that, in the absence of lipoproteins, TAG would exist in aqueous blood as immiscible oil droplets, while free fatty acids liberated from TAG and phospholipids in the absence of the blood protein albumin would act as detergents and dissolve cell membranes.
Lipoprotein structure: a shopping bag and groceries
The general structure of a lipoprotein consists of a central core of hydrophobic, neutral lipid (TAG and CE) surrounded by a hydrophilic coat of phospholip-ids, free cholesterol, and apoproteins. A useful analogy for this arrangement of molecules is that of a “shop-ping bag and groceries,” with the lipid core represent-ing the groceries and the outer coat the fabric of the bag. The apoproteins weave in and out of the lipid core and outer surface layer and form the thread of the fabric which holds the bag together (see Figure 6.5). This clever arrangement of molecules renders the hydrophobic lipids soluble for the purpose of transport in blood. In addition to conferring struc-tural integrity on the lipoprotein particle, apoproteins have a vital role in controlling the metabolism of lipoproteins by acting as ligands for cell membrane receptors and cofactors for key enzymes.
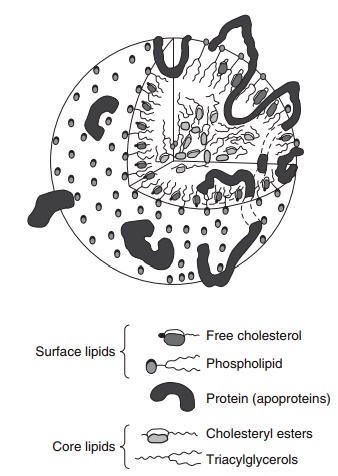
Figure 6.5 General lipoprotein structure. (Reproduced from Dur-rington PN. Hyperlipidaemia Diagnosis and Management, 2nd edn. Elsevier Science, Oxford, copyright 1995 with permission of Elsevier.)
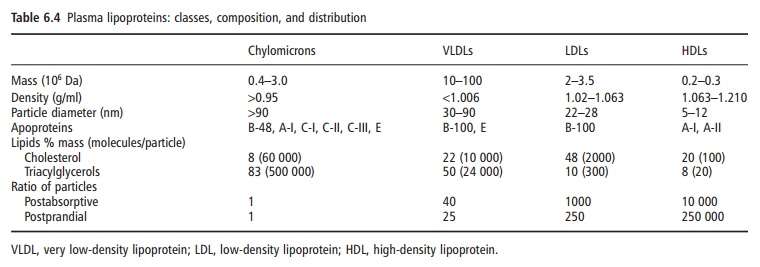
Plasma lipoproteins can be subdivided into distinct classes on the basis of their physical properties and/or composition, both of which reflect the physiological role in the transport of lipids from sites of synthesis (endogenous lipids) and absorption (exogenous lipids, absorbed in the gut) to sites of storage (adipose tissue) and utilization (skeletal muscle). The principal classes of lipoproteins are traditionally defined by density, which is determined by the ratio of lipid to protein in the lipoprotein particle. Since lipids tend to occupy a greater molecular volume than proteins, they are lighter and less dense. Thus, particles with high lipid content are larger and less dense (carry more lipid groceries) than lipoproteins enriched with protein. This property relates directly to the transport function and metabolic interrelationships between lipoprotein classes in blood. It can also be used to separate lipoproteins of different densities because lipoproteins of different density have different flota-tion characteristics in the ultracentrifuge (note that plasma lipoproteins will float when subjected to cen-trifugal force, whereas pure proteins sink). Other clas-sification schemes for plasma lipoproteins have exploited differences in their net electrical charge (electrophoretic mobility), particle size (exclusion chromatography, gradient gel electrophoresis), and immunological characteristics conferred upon the lipoprotein by the types of apoproteins in each lipoprotein subclass (see Table 6.4). Some of these techniques permit the further resolution of VLDL, low-density lipoproteins (LDLs), and high-density lipoproteins (HDLs) into discrete subclasses, the dis-tribution of which relates to cardiovascular risk and is determined by genetic and lifestyle factors.
Related Topics