Chapter: Medical Immunology: The Induction of an Immune Response: Antigens, Lymphocytes, and Accessory Cells
Induction of the Immune Response
INDUCTION OF THE IMMUNE RESPONSE
A. Immune Recognition: Clonal Restriction and Expansion
For the initiation of an immune response, an antigen or a peptide associated with a MHC molecule must be recognized as nonself by the immunocompetent cells. This phenomenon is designated immune recognition. The immune system of a normal individual may recog- nize as many as 106–108 different antigenic specificities. An equal number of different small families (clones) of lymphocytes, bearing receptors for the different antigens, con-stitute the normal repertoire of the immune system. Each immunocompetent cell expresses on its membrane many identical copies of a receptor for one single antigen. Thus, a major characteristic of the immune response is its clonal restriction, i.e., one given epitope will be recognized by a single family of cells with identical antigen receptors, known as a clone. When stimulated by the appropriate specific antigen, each cell will proliferate, and the clone of reactive cells will become more numerous (clonal expansion).
Since most immunogens present many different epitopes to the immune system, the normal immune responses are polyclonal, i.e., multiple clones of immunocompetent cells, each one of them specific for one unique epitope, are stimulated by any complex im-munogen.
B. The Antigen Receptor on T and B Lymphocytes
In B lymphocytes, the antigen receptors are membrane-inserted immunoglobulins, partic-ularly IgD and monomeric IgM molecules . In T lymphocytes the antigen receptors are known as T-cell receptors (TcR).
Two types of TcR have been identified, depend-ing on the polypeptide chains that constitute them. In differentiated T cells, the TcR most frequently found is constituted by two polypeptide chains, designated as α and β (αβ TcR), with similar molecular weights (40,000–45,000). A second type of TcR, constituted by two different polypeptide chains known as γ and δ (γδ TcR), is predominantly found in the sub-mucosal lymphoid tissues.
The two chains of the αβ TcR have extracellular segments with variable and constant domains, short cytoplasmic domains, and a transmembrane segment (Fig. 4.5). A disulfide bridge joins them just outside the transmembrane segment.
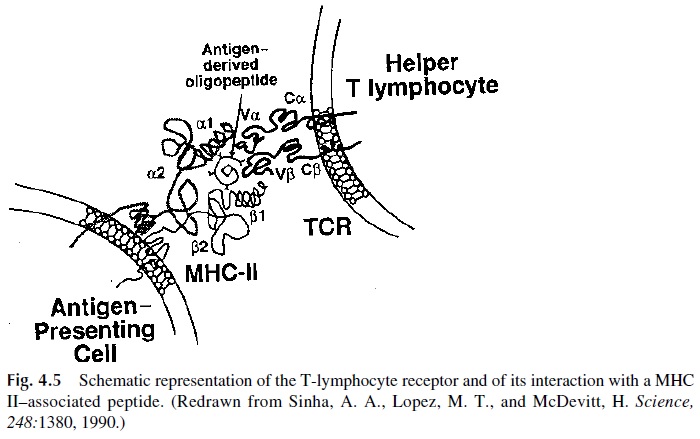
The β chains are highly polymorphic and are encoded by a multigene family that in-cludes genes for regions homologous to the V, C, D, and J regions of human immunoglobulins.
The chains are encoded by a more limited multigene family with genes for regions homologous to the V, C, and J regions of human immunoglobulins . Simi- lar polymorphisms have been defined for the γδ chains. Together, the variable regions of αβ and γδchains define the specific binding sites for peptide epitopes presented in associ- ation with MHC-II molecules.
C. Antigen Processing and Presentation
Most immune responses to complex, T-dependent antigens involve the participation of sev-eral cell types, including T and B lymphocytes that are directly involved in the generation of effector mechanisms and accessory cells that assist in the inductive stages of the immune response. Antigen-presenting cells (APCs) are accessory cells that express MHC-II molecules on their membrane where antigen fragments can be bound and “presented” to lymphocytes. Additionally, they often express ligands for costimulatory molecules and re-lease cytokines that assist the proliferation and/or differentiation of T and B lymphocytes. Several types of cells can function as APCs. The most effective are the dendritic cells found in the paracortical areas of the lymph nodes. Dendritic cells and their precursors (Langerhans cells in the dermis) as well as activated B cells serve as effective APCs in immunologically naïve hosts. Tissue macrophages and related cells are effective APCs in that they have phagocytic properties and can process proteins and present the resulting peptides on MHC-II molecules. Macrophages become particularly effective when antigen molecules are coated with IgG and complement, under which conditions their phagocytic potential is fully expressed.
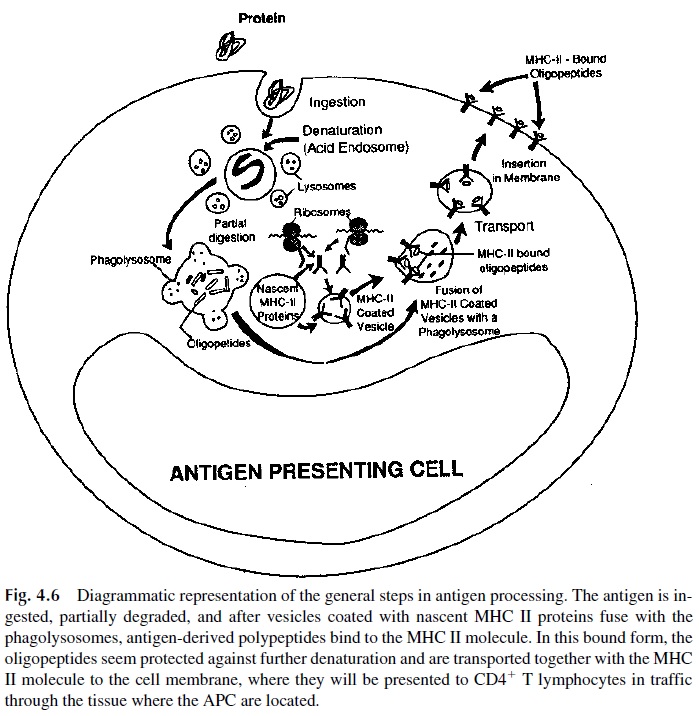
Antigen processing and presentation is a complex sequence of events that involves endocytosis of antigens on membrane patches and transport to an acidic compartment (lysosome) within the cell that allows antigen degradation into small peptides. As antigens are broken down, vesicles coated with newly synthesized HLA II molecules fuse with the lysosome. Some of the peptides generated during processing have affinity for the binding site located within the MHC-II heterodimer. The resulting MHC-peptide complexes are then transported to the APC cell membrane, where they can interact with and activate T cells bearing the T-cell receptor specific for the peptide (Fig. 4.6).
D. Activation of Helper T Lymphocytes
The activation of resting T helper cells requires a complex and coordinated sequence of sig-nals delivered from the T-cell receptor on the cell membrane to the nucleus of the cell. Of all the signals involved, the only antigen-specific interaction is the one that involves the TcR and the peptide-MHC complex. The binding of the peptide-MHC complex to the TcR is of low affinity, and other receptor-ligand interactions are required to maintain T-lym-phocyte adhesion to APC and for the delivery of required secondary signals.
The TcR on a helper T lymphocyte interacts with both the antigen-derived peptide and the MHC-II molecule (Fig. 4.5). This selectivity of the TcR from helper T lymphocytes to interact with MHC-II molecules results from selection in the thymus. During thymic on-togeny, the differentiation of helper and cytotoxic T lymphocytes is based on the ability of their TCR to interact with MHC-II molecules (helper T lymphocytes) or with MHC-I molecules (cytotoxic T lymphocytes) . The interactions between T lym-phocytes and MHC-expressing cells are strengthened by cell surface molecules, which also interact with constant (not peptide-loaded) regions of MHC molecules: the CD4 molecule on helper T cells interacts with MHC-II molecules, while the CD8 molecule on cytotoxic lymphocytes interacts with MHC-I molecules.
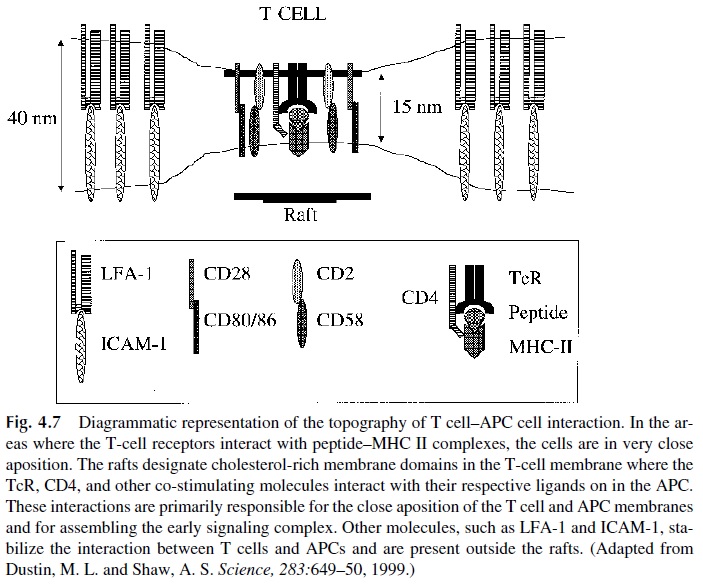
Several other cell adhesion molecules (CAM) can mediate lymphocyte-AC interac-tions, including lymphocyte function–associated antigen (LFA)-1 interacting with the in-tercellular adhesion molecules (ICAM)-1, -2, and -3, CD2 interacting with CD58 (LFA-3). Unlike the interactions involving the TcR, these interactions are not antigen specific. Their role is to promote stable adhesion and signaling between T lymphocytes and APC essential for proper stimulation through the TcR. Furthermore, T-cell activation requires sustained signaling achieved through the establishment of what is known as the immunological synapse, in which peptide–MHC-II complexes form clusters on the APC membrane allow-ing aggregation and clustering of multiple TcR molecules on the opposing T-cell membrane. In the regions of contact the two cells are separated by a narrow gap surrounded by other in-teracting molecules such as (CD2/CD58, LFA-1/ICAM-1, etc.). The result is a stable and close apposition between APC and T cell, essential for sustained signaling (Fig. 4.7).
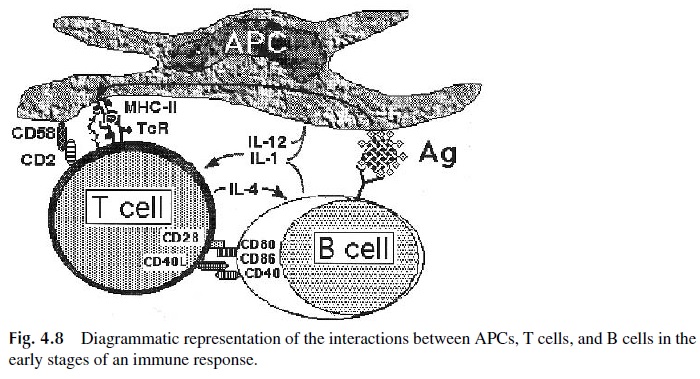
It is important to stress that accessory cells participate in the activation of helper T lymphocytes through the delivery of signals involving cell-cell contact as well as by the release of soluble factors or cytokines (Fig. 4.8):
1. Signals mediated by CD4–MHC-II interactions
2. Signals mediated by the cell-cell interactions, which are facilitated by the up-regulation of some of the interacting molecules after initial activation, includ-ing:
CD2 (T cell): CD58 (APC)
LFA-1 (T cell): ICAM-1, ICAM-2, ICAM-3 (APC) CD40 L (T cell): CD40 (APC)
CD28 (T cells): CD80, CD86 (APC)
3. Signals mediated by interleukins:
Interleukin-1 (IL-1), a cytokine that promotes growth and differentiation of many cell types, including T and B lymphocytes. Both membrane-bound and soluble IL-1 have been shown to be important in activating T lymphocytes in vitro. Mem-brane-bound IL-1 can only activate T lymphocytes in close contact with the APC.
Interleukin-12 (IL-12) promotes Th1 cell differentiation.
However, all these co-stimulatory signals are not specific for any given antigen. The speci-ficity of the immune response is derived from the essential and first activation signal de-livered through the antigen-specific TcR.
The precise sequence of intracellular events resulting in T-cell proliferation and dif-ferentiation. The following is a summary of the major steps in the activation sequence:
1.Occupancy and cross-linking of the TcR signals the cell through a closely as-sociated complex of molecules, known as CD3, that has signal-transducing properties. The TcR heterodimer itself has no recognizable kinase activity. The associated CD3 complex, however, has 10 intracytoplasmic motifs (im-munoreceptor-tyrosine-based activation motifs, ITAMs) that play a key role in the sequence of cell activation. These ITAMs are associated with the γ, δ, ε, and ζ chains of the CD3 complex.
2.Co-stimulatory signals are delivered by CD4, as a consequence of the interac-tion with MHC-II, and by CD45, a tyrosine phosphatase, activated as a conse-quence of TcR occupancy.
3.The activation of CD45 initiates the sequential activation of several protein ki-nases closely associated with CD3 and CD4. The activation of the kinase cas-cade has several effects, namely:
a. Phospholipase C activation, leading to the mobilization of Ca2+ -depen-dent second messenger systems, such as inositol triphosphate (IP3), which promotes an increase in intracellular free Ca2+ released from intracellular organelles and taken up through the cell membrane. The increase in intra-cellular free calcium results in activation of a serine threonine phosphatase known as calcineurin. Diacylglycerol (DAG), another product released by phospholipase C, activates another serine/threonine kinase known as pro-tein kinase C (PKC). Multiple other enzymes and adapter molecules are activated in the ensuing cascading sequence.
b. The activation of second messenger systems results in the activation and translocation of transcription factors, such as the nuclear factor-kappa B (NF- B) and the nuclear factor of activated T cells (NF-AT). Once translo-cated to the nucleus, these factors induce genes controlling cytokine production and T-cell proliferation, such as those encoding interleukin-2 (IL-2), the IL-2 receptor, and c-myc.
c. Upregulation and modification of several membrane proteins on the T-cell membrane, such as CD28 and CD40 ligand (CD40L).These molecules in-teract, respectively, with CD80/86 and CD40 on the APC membrane. The interactions involving this second set of molecules deliver additional sig-nals that determine the continuing proliferation and differentiation of anti-gen-stimulated T cells.
E. Antigen Presentation and Activation of Cytotoxic T Lymphocytes
If an APC is infected by an intracellular organism (virus, bacteria, or parasite), the infect-ing agent will multiply in the cytosol. Peptides derived from mi-crobial proteins are loaded onto MHC-I molecules, transported to the cell surface, and pre-sented to cytotoxic T lymphocytes (CTL). CTL are a special population of effector T cells capable of killing target cells bearing specific antigen and are largely CD8+ .
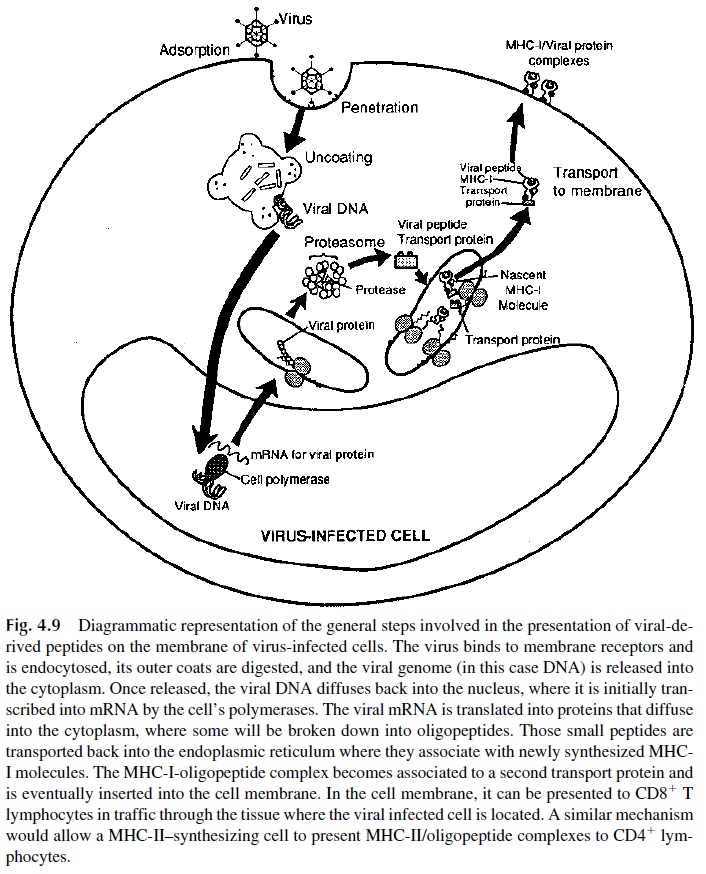
The way in which MHC I molecules and viral peptides become associated has re-cently been elucidated (Fig. 4.9). Upon infection, intracellular microbes start to produce their own proteins. Some of the nascent microbial proteins diffuse into the cytoplasm, where they become associated with degradative enzymes forming a peptide-enzyme com-plex (proteasome). In these complexes, the protein is partially digested, and the resulting peptides bind to specialized proteins (transporters associated with antigen processing, TAP), which deliver them to the endoplasmic reticulum, the site of MHC-I synthesis and assembly. In the endoplasmic reticulum, the viral peptides bind to newly synthesized MHC class I molecules, and the resulting MHC–viral peptide complex is transported to the mem-brane of the infected cell.
Similar to helper T cells, the stimulation of cytotoxic T cells also requires additional signals and interactions, some of which depend upon cell-cell contact, such as those medi-ated by the interaction of CD8 with MHC I, CD2 with CD58, LFA-1 with ICAM family members, and CD28 with CD80 and CD86. On the other hand, the expansion of antigen-activated cytotoxic T lymphocytes requires the secretion of IL-2. Rarely, activated cyto-toxic T lymphocytes can secrete sufficient quantities of IL-2 to support their proliferation and differentiation and thus proceed without help from other T-cell subpopulations. Most frequently, activated helper T lymphocytes provide the IL-2 necessary for cytotoxic T lym-phocyte differentiation. The activation of helper T cells requires the presentation of anti-gen-derived peptides in association with MHC-II molecules. The simultaneous expression of immunogenic peptides in association with MHC-I and MHC-II molecules is likely to happen in infected APCs.
Resting, circulating cytotoxic T lymphocytes carry antigen receptors able to recog-nize associations of MHC-I and nonself peptides; occupancy of the binding site on the TCR by MHC-I–associated peptide provides the antigen-specific signal that drives cytotoxic T cells. In the case of antiviral responses, virus-infected macrophages are also likely to ex-press viral peptide–MHC-II complexes on their membrane; these complexes are able to ac-tivate CD4+ helper T cells. Thus, an infected professional APC is likely to fulfill the con-ditions required to serve as the anchor for all the cells participating in the immune response against the infecting virus.
Cytotoxic T lymphocytes also differentiate and proliferate when exposed to cells from an individual of the same species but from a different genetic background, as a consequence of tissue or organ transplantation.
In vitro, the degree of allostimulation between lymphocytes of two different individuals can be assessed by the mixed lymphocyte reac-tion . Two types of recognition have been analyzed:
1. Donor peptides presented by nonpolymorphic MHC molecules of donor cells appear to trigger the initial stages of the mixed lymphocyte reaction.
2. As a consequence of the release of cytokines during the initial activation of the rejection reaction, the expression of nonself MHC molecules is upregulated on donor tissues. Donor MHC molecules are shed into the circulation, taken up by professional APCs of the recipient, and processed and presented as nonself pep-tides associated with self MHC molecules to the immune system of the recipient.
MHC-II–expressing cells must be present for the mixed lymphocyte reaction to take place. This requirement suggests that activation of helper T cells by recognition of MHC-II–peptide complexes is essential for the differentiation of cytotoxic CD8+ cells. The role of helper T cells in the mixed lymphocyte reaction is likely to be very similar to the role of helper T cells that assist B-cell responses, i.e., to provide cytokines and co-stimulatory sig-nals essential for cytotoxic T-cell growth and differentiation.
Graft rejection is more intense with increasing MHC disparity between donor and host. This is likely a consequence of the greater structural differences between the non-shared MHC molecules. Thus, MHC differences are likely to perpetuate the rejection reac-tion in that self (host-derived) MHC molecules will present nonself peptides derived from the donor’s MHC molecules.
Alternative pathways of antigen presentation to cytotoxic T cells involving nonpoly-morphic molecules have been recently described. The best characterized of these pathways involves CD1, a family of nonpolymorphic, MHC-I–related molecules that includes five different isomorphic forms (A to E). Antigen-presenting cells, including dendritic cells and B cells, express CD1A, B, and C molecules and have been shown to present mycobac-terium-derived lipid and lipoglycan antigens to both γδ and αβ CD8+ cytotoxic T lym-phocytes. Both γδ and CD8+ αβ T cells stimulated by mycobacterial antigens presented in association with CD1 molecules cause the death of the presenting cells.
Related Topics