Chapter: Biology of Disease: Disorders of the Gastrointestinal Tract, Pancreas, Liver and Gall Bladder
Gastrointestinal Tract and its Accessory Organs
THE GASTROINTESTINAL TRACT AND
ITS ACCESSORY ORGANS
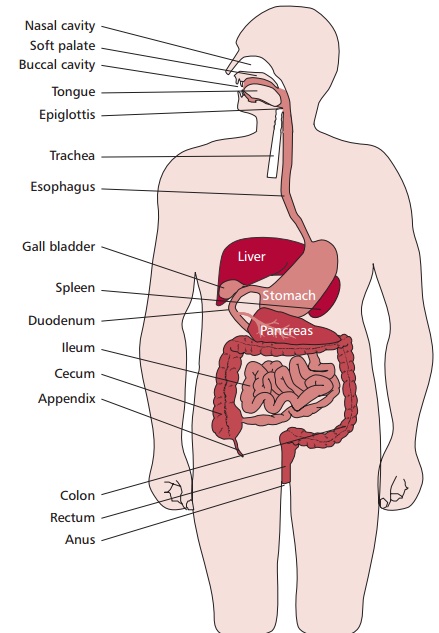
The GIT may be described as a tubular system with
distinctive regions that begin with the mouth leading to the pharynx,
esophagus, stomach, the small and large intestines and terminating at the anus
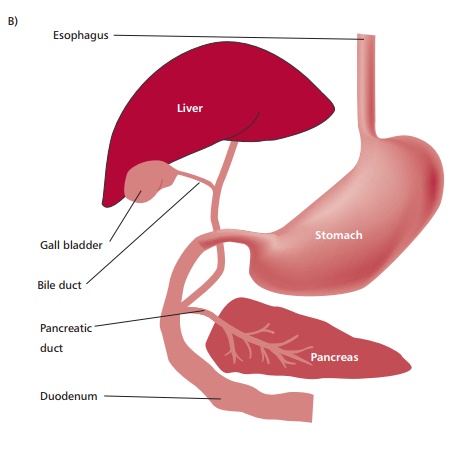
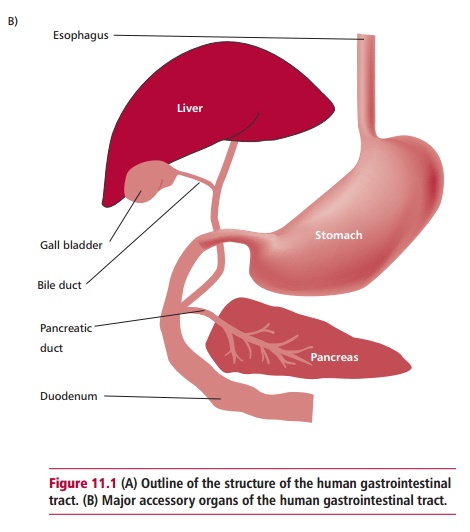
(Figure 11.1 (A)). It is associated with several accessory digestive organs, such
as the pancreas, liver and gall bladder (Figure
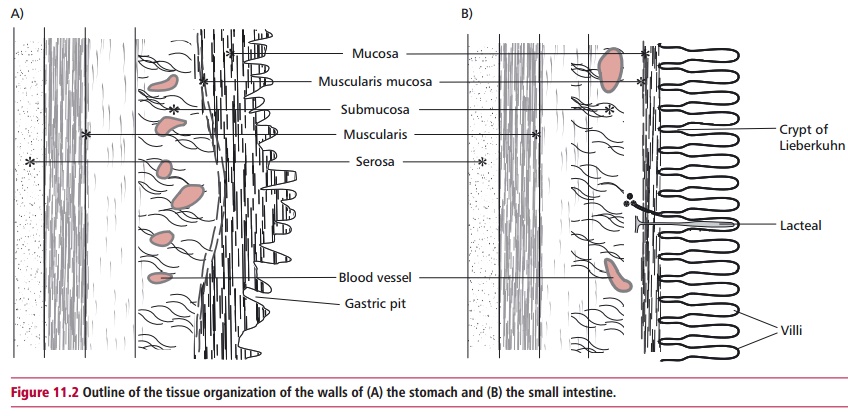
11.1 (B)). The walls of the GIT
have a fairly common substructure (Figure
11.2 (A) and (B)) and its interior is called the
lumen. The GIT and accessory organs are bound to each other and to the inner
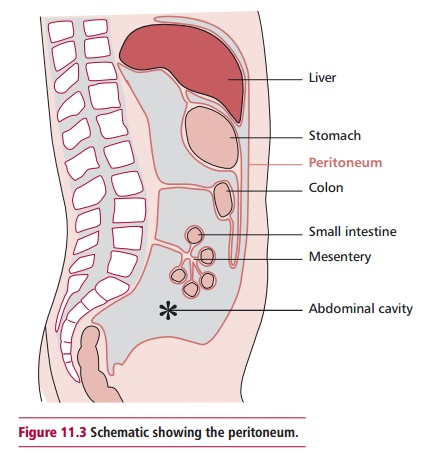
wall of the abdomen by the peritoneum.
This is a strong, colorless membrane with a smooth surface that consists of two
parts; the parietal peritoneum, which lines the abdominal cavity, and the
visceral peritoneum that covers most of the organs in the abdomen (Figure 11.3). The thin space between the
two parts, called the peritoneal cavity, is filled with serous fluid. In males,
the peritoneum forms a closed sac but in females it is con-tinuous with the
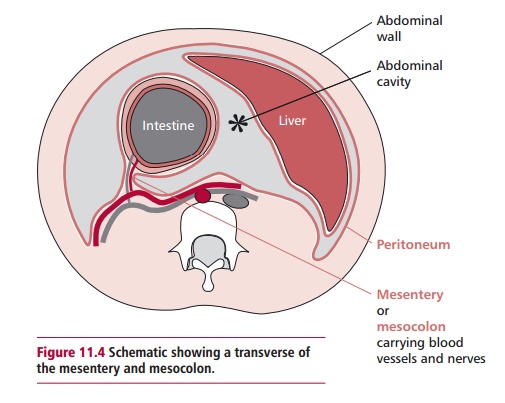
mucous membrane of the uterine tubes. The mesentery
is a fan shaped portion of peritoneum that extends from the posterior
abdom-inal wall and wraps around the small intestine and returns to its origin
forming a double layer (Figure 11.4).
The mesentery contains the blood and
lymphatic vessels and nerves that supply the small
intestine. Another peri-toneal fold, the mesocolon
(Figure 11.4), has similar functions
with respect to the large intestine. The mesentery and mesocolon anchor the
intestines in place but only loosely, allowing them considerable movement as
muscu-lar activities mix and move the contents through the GIT.
The pancreas is found near the stomach and small
intestine (Figure 11.1 (A) and (B)) and functions as both an exocrine and an endocrine gland. In
its exo-crine role it produces pancreatic juice that is released into the first
part of the small intestine, the duodenum, through the pancreatic duct. This
secretion contains precursors of digestive enzymes that are activated and
function in the GIT, and also hydrogen carbonate (HCO3–) that
increases the pH of GIT contents. This latter function is vital since the
material leaving the stomach is acidic. The pancreas produces about 1200–1500
cm3 of pancreatic juice per
day. Within the tissues of the pancreas are groups of
cells called the islets of Langerhans. The cells in the islets have endocrine
functions and produce and secrete the peptide hormones glucagon and insulin that
regulate the concen-tration of glucose in the blood .
The liver is the largest organ in the body (Figure 11.1 (A) and (B)), weighing
about 1.5 kg. It has two major lobes, each divided into functional units called
lobules. Lobules consist of cuboidal cells arranged around a central vein into
which the blood drains. Between the plates of hepatic cells are sinusoids that
are supplied by the portal vein and hepatic artery. The sinusoids are lined
with phagocytic Kupffer cells that belong to the reticuloendothelial system and
their main function is to engulf bacteria and other foreign particles in blood.
Thus there is a dual blood supply to liver, with blood coming from the
digestive tract and spleen through the portal vein, and from the aorta in the
hepatic artery (Figure 14.2). About a
third of incoming blood is arterial, and brings oxygen, whereas two thirds is
venous from the portal vein.
The large reserve capacity of the liver means that it
needs only 10–20% of its tissues to be functioning to sustain life. It also has
a remarkable ability to regenerate itself after its tissue has been removed or
destroyed by dis-ease. Complete destruction or removal of the liver results in
death within 10 h, hence liver disease with loss of function is a serious
matter. The liver has numerous functions. It acts as an interface between
theGIT and the rest of the body tissues because the hepatic portal vein
car-ries blood directly to it from the GIT. Hence it is able to regulate the
post-hepatic blood concentrations of many of the nutrients absorbed by the GIT.
Similarly, the liver also regulates the concentrations of many biomolecules
produced by the body, for example steroid hormones, and deals with many toxins,
such as drugs, pesticides and carcinogens, to render them less harm-ful and
solubilizes them for excretion . The liver also produces many of the plasma
proteins including albumin and the clotting factors . Bile produced by the
hepatocytes is secreted into bile canal-iculi and eventually drains into the
bile duct. About 700–1200 cm3 of bile are
produced daily and stored and concentrated in a hollow organ called the gall
bladder (Figure 11.1 (A) and (B)) prior to its release in the small intestine. Bile does not
contain enzymes, but it does contain bile salts, for example sodium
glycocholate and taurocholate. These are detergents that aid in the digestion
of lipids by emulsifying them to form water-soluble complexes.
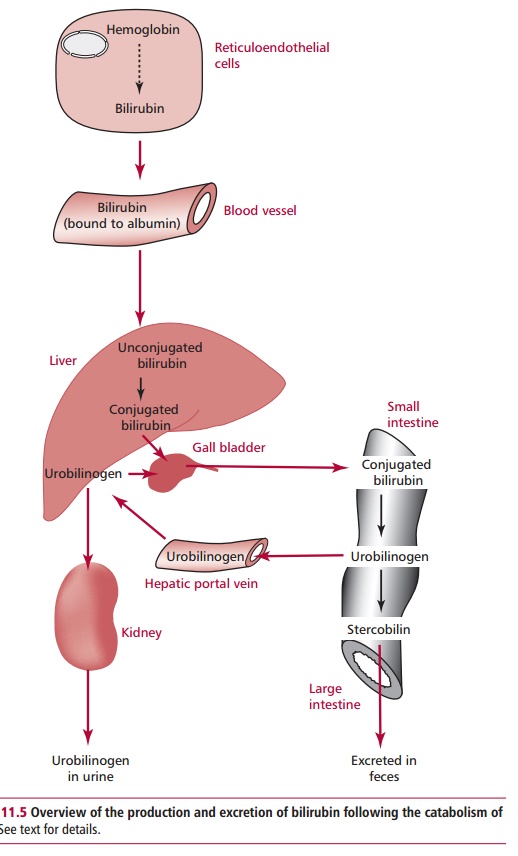
Bile pigments are derived from heme which results
from the destruction of old erythrocytes . In the spleen (Figure 11.1 (A) and (B)), destruction of red blood cells
releases hemoglobin, which is then cata-bolized to free heme and globin. The
latter is degraded to amino acids. Bilirubin is derived from heme, the
iron-containing protoporphyrin ring of hemoglobin. A typical adult produces
around 450 µmol of bilirubin per day. This bilirubin is referred to as
unconjugated bilirubin. It is insoluble in water and is transported in the
plasma bound to albumin to be taken up by hepatocytes. Here, it is conjugated
with glucuronic acid by UDP-glucuronyltransferase to form mono- and
diglucuronides in a manner resembling the detoxification reactions described.
Conjugated bilirubin is much more water soluble than its unconjugated form and
is secreted into the bile duct. In the small intestine, conjugated bilirubin is
a substrate for bacteria that convert it to urobilinogen and stercobilin. Most
stercobilin is excreted in feces, although some urobilinogen is absorbed and
taken to the liver in the hepatic portal vein and re-excreted in bile or by the
kidneys (Figure 11.5).
Related Topics