Chapter: Medical Physiology: Pregnancy and Lactation
Function of the Placenta: Developmental and Physiologic Anatomy of the Placenta
Function of the Placenta
Developmental and Physiologic Anatomy of the Placenta
While the trophoblastic cords from the blastocyst are attaching to the uterus, blood capillaries grow into the cords from the vascular system of the newly forming embryo. By the 16th day after fertilization, blood also begins to be pumped by the heart of the embryo itself. Simultaneously, blood sinuses supplied with blood from the mother develop around the outsides of the tro-phoblastic cords. The trophoblast cells send out more and more projections, which become placental villi into which fetal capillaries grow. Thus, the villi, carrying fetal blood, are surrounded by sinuses that contain maternal blood.
The final structure of the placenta is shown in Figure 82–5. Note that the fetus’s blood flows through two umbilical arteries, then into the capillaries of the villi,and finally back through a single umbilical vein into the fetus. At the same time, the mother’s blood flows from her uterine arteries into largematernal sinuses that sur-round the villi and then back into the uterine veins of the mother. The lower part of Figure 82–5 shows the relation between the fetal blood of each fetal placental villus and the blood of the mother surrounding the out-sides of the villus in the fully developed placenta.
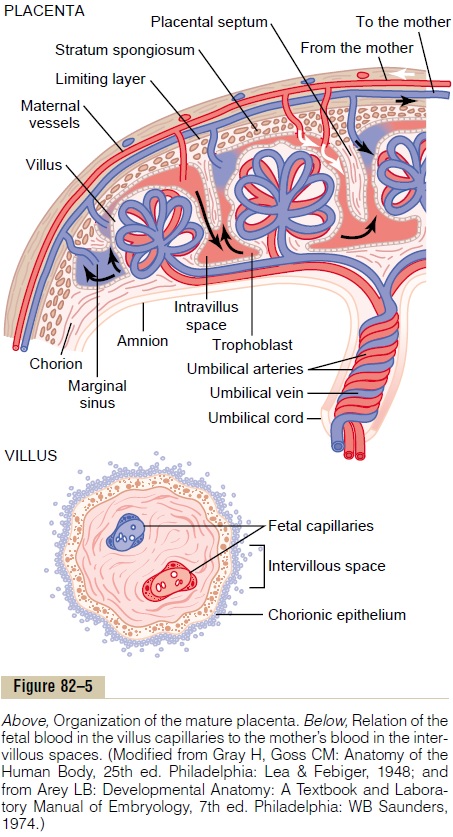
The total surface area of all the villi of the mature pla-centa is only a few square meters—many times less than the area of the pulmonary membrane in the lungs. Nev-ertheless, nutrients and other substances pass through this placental membrane mainly by diffusion in much the same manner that diffusion occurs through the alveolar membranes of the lungs and the capillary membranes elsewhere in the body.
Placental Permeability and Membrane Diffusion Conductance
The major function of the placenta is to provide for diffusion of foodstuffs and oxygen from the mother’s blood into the fetus’s blood and diffusion of excretoryproducts from the fetus back into the mother.
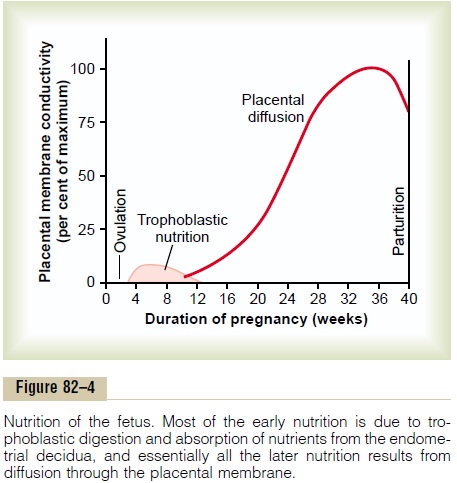
In the early months of pregnancy, the placental membrane is still thick because it is not fully devel- oped. Therefore, its permeability is low. Further, the surface area is small because the placenta has not grown significantly. Therefore, the total diffusion con- ductance is minuscule at first. Conversely, in later preg- nancy, the permeability increases because of thinning of the membrane diffusion layers and because the surface area expands many times over, thus giving the tremendous increase in placental diffusion shown in Figure 82–4.
Rarely, “breaks” occur in the placental membrane, which allows fetal blood cells to pass into the mother or, even less commonly, the mother’s cells to pass into the fetus. Fortunately, it is rare for the fetus to bleed severely into the mother’s circulation because of a rup- tured placental membrane.
Diffusion of Oxygen Through the Placental Membrane.
Almost the same principles for diffusion of oxygen through the pulmonary membrane are applicable for diffusion of oxy-gen through the placental membrane. The dissolved oxygen in the blood of the large maternal sinuses passes into the fetal blood by simple diffusion, driven by an oxygen pressure gradient from the mother’s blood to the fetus’s blood. Near the end of pregnancy, the mean PO2 of the mother’s blood in the placental sinuses is about 50 mm Hg, and the mean PO2 in the fetal blood after it becomes oxygenated in the placenta is about 30 mm Hg. Therefore, the mean pressure gra-dient for diffusion of oxygen through the placental membrane is about 20 mm Hg.
One might wonder how it is possible for a fetus to obtain sufficient oxygen when the fetal blood leaving the placenta has a PO2 of only 30 mm Hg. There are three reasons why even this low PO2 is capable of allowing the fetal blood to transport almost as much oxygen to the fetal tissues as is transported by the mother’s blood to her tissues.
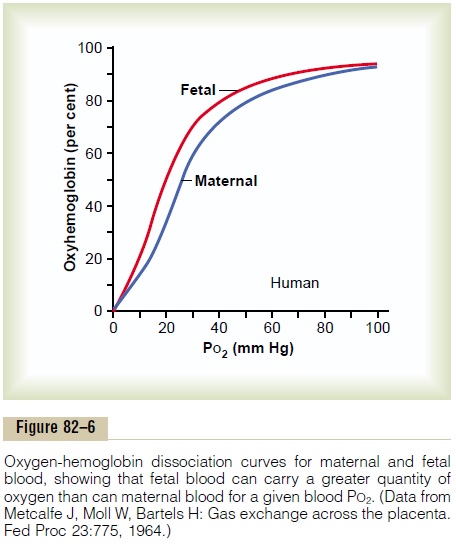
First, the hemoglobin of the fetus is mainly fetalhemoglobin, a type of hemoglobin synthesized in thefetus before birth. Figure 82–6 shows the comparative oxygen dissociation curves for maternal hemoglobin and fetal hemoglobin, demonstrating that the curve for fetal hemoglobin is shifted to the left of that for mater-nal hemoglobin. This means that at the low PO2 levels in fetal blood, the fetal hemoglobin can carry 20 to 50 per cent more oxygen than maternal hemoglobin can.
Second, the hemoglobin concentration of fetal bloodis about 50 per cent greater than that of the mother; thisis an even more important factor in enhancing the amount of oxygen transported to the fetal tissues.
Third, the Bohr effect, which is explained in relation to the exchange of carbon dioxide and oxygen in the lung, provides another mechanism to enhance the transport of oxygen by fetal blood.
That is, hemoglobin can carry more oxygen at a low PCO2 than it can at a high PCO2. The fetal blood entering the placenta carries large amounts of carbon dioxide, but much of this carbon dioxide diffuses from the fetal blood into the maternal blood. Loss of the carbon dioxide makes the fetal blood more alkaline, whereas the increased carbon dioxide in the maternal blood makes it more acidic.
These changes cause the capacity of fetal blood to combine with oxygen to increase and that of maternal blood to decrease. This forces still more oxygen from the maternal blood, while enhancing oxygen uptake by the fetal blood. Thus, the Bohr shift operates in one direction in the maternal blood and in the other direc-tion in the fetal blood. These two effects make the Bohr shift twice as important here as it is for oxygen exchange in the lungs; therefore, it is called the doubleBohr effect.
By these three means, the fetus is capable of receiv-ing more than adequate oxygen through the placental membrane, despite the fact that the fetal blood leaving the placenta has a PO2 of only 30 mm Hg.
The total diffusing capacity of the entire placenta for oxygen at term is about 1.2 milliliters of oxygen per minute per millimeter of mercury oxygen pressure dif-ference across the membrane.This compares favorably with that of the lungs of the newborn baby.
Diffusion of Carbon Dioxide Through the Placental Membrane.
Carbon dioxide is continually formed in the tissues of the fetus in the same way that it is formed in maternal tissues, and the only means for excreting the carbon dioxide from the fetus is through the placenta into the mother’s blood. The PCO2 of the fetal blood is 2 to 3 mm Hg higher than that of the maternal blood. This small pressure gradient for carbon dioxide across the membrane is more than sufficient to allow adequate diffusion of carbon dioxide, because the extreme sol-ubility of carbon dioxide in the placental membrane allows carbon dioxide to diffuse about 20 times as rapidly as oxygen.
Diffusion of Foodstuffs Through the Placental Membrane.
Other metabolic substrates needed by the fetus diffuse into the fetal blood in the same manner as oxygen does. For instance, in the late stages of pregnancy, the fetus often uses as much glucose as the entire body of the mother uses. To provide this much glucose, the trophoblast cells lining the placen-tal villi provide for facilitated diffusion of glucose through the placental membrane. That is, the glucose is transported by carrier molecules in the trophoblast cells of the membrane. Even so, the glucose level in fetal blood is 20 to 30 per cent lower than that in maternal blood.
Because of the high solubility of fatty acids in cell membranes, these also diffuse from the maternal blood into the fetal blood, but more slowly than glucose, so that glucose is used more easily by the fetus for nutrition. Also, such substances as ketone bodies and potassium, sodium, and chloride ions diffuse with relative ease from the maternal blood into the fetal blood.
Excretion of Waste Products Through the Placental Membrane.
In the same manner that carbon dioxide diffuses from the fetal blood into the maternal blood, other excretory products formed in the fetus also diffuse through the placental membrane into the maternal blood and are then excreted along with the excretory products of the mother. These include especially the nonprotein nitrogens such as urea, uric acid, and crea-tinine. The level of urea in fetal blood is only slightlygreater than that in maternal blood, because urea dif-fuses through the placental membrane with great ease.
However, creatinine, which does not diffuse as easily, has a fetal blood concentration considerably higher than that in the mother’s blood. Therefore, excretion from the fetus occurs mainly, if not entirely, as a result of diffusion gradients across the placental membrane, because there are higher concentrations of the excre-tory products in the fetal blood than in the maternal blood.
Related Topics