Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Communities, ecosystems, and the functional role of fishes
Fishes in food webs
Fishes in food webs
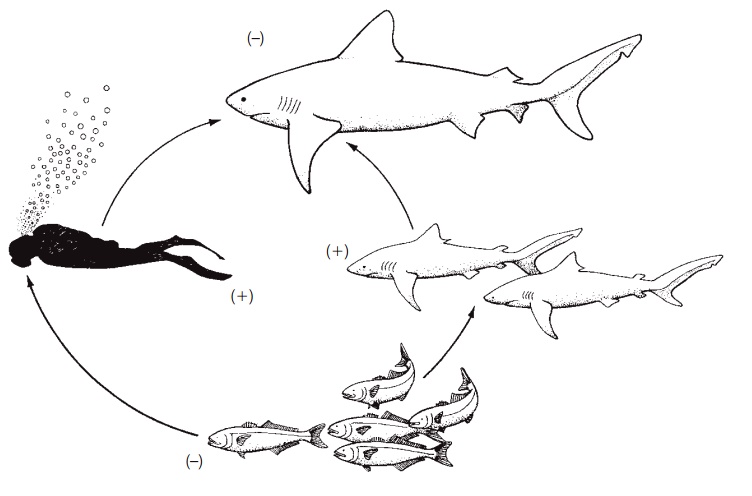
Discussing top-down and bottom-up interactions can imply that linear food chains characterize many aquatic ecosystems. This is, however, a simplifi cation of a much more dynamic situation because few straight line chains (“A eats B who eats C who eats D”) exist. Instead, trophic interactions often involve herbivores that eat animals, carnivores that eat plants, cannibalism, reversals in energy transfer (one species eats juveniles of another species but winds up as the prey of the adults of the same species), and fishes that parasitize their predators by eating their fins, scales, and mucus. Most individuals enter a detrital food loop, either directly during decomposition or indirectly after falling prey and being processed into feces. Hence trophic interactions are more accurately described as food webs than food chains, and when disrupted can cause surprising results (Polis et al. 2004; see Fig. 25.14).
A particularly thorough analysis of the role of fishes in a community food web has been conducted for lowland streams in Venezuela and Costa Rica (Winemiller 1990b). The fish assemblages at four different sites included between 20 and 83 species that fed on detritus, plants, seeds, flowers, protozoans, aquatic and terrestrial insects, numerous aquatic invertebrates (worms, crabs, shrimps, clams, snails), fishes, larval amphibians, turtles, lizards, birds, and mammals. Food web analysis of relationships only between fish and their prey indicated between 50 and 100 interacting taxa and 200–1200 trophic links (i.e., one fish species might eat a dozen different food types, each one constituting a link) (Fig. 25.16). The pimelodid catfish Rhamdia fed at three different trophic levels: seeds, prawns, and fishes. More than a dozen species (mostly characins, catfishes, and cichlids) were omnivores, feeding extensively on both plant and animal matter. Detritus, formed from decaying aquatic and terrestrial vegetation, was a particularly important component, accounting for 30–50% of the food eaten by all species. Species as diverse as characins, catfishes, livebearers, cichlids, and sleepers fed directly on detritus.
Reciprocal food loops were common. The trichomycterid catfish Ochmacanthus fed on the external mucus of Oscars (Astronotus ocellaris); Oscars in turn ate the catfish. A predatory cichlid (Cichlasoma dovii) ate juvenile Sleepers (Gobiomorus dormitor), while adult Sleepers ate juvenile cichlids.
As complex as the interactions in Fig. 25.16 appear, the actual food web is even more intricate. Different ontogenetic stages of a species were not separated in the analysis; nine dominant piscivorous species fed initially on zooplankton, switching later to invertebrates and eventually to fishes as they grew larger. The food webs at the four sites differed considerably due to varying diversities and species compositions, but differences also occurred within sites during the wet versus dry season. Even this seemingly complex description of interactions simplifi es the true complexity of feeding relationships in a natural community, because only the interactions involving fishes are listed; other food items of the prey of the fishes were not considered (i.e., links between shrimp and snails or between
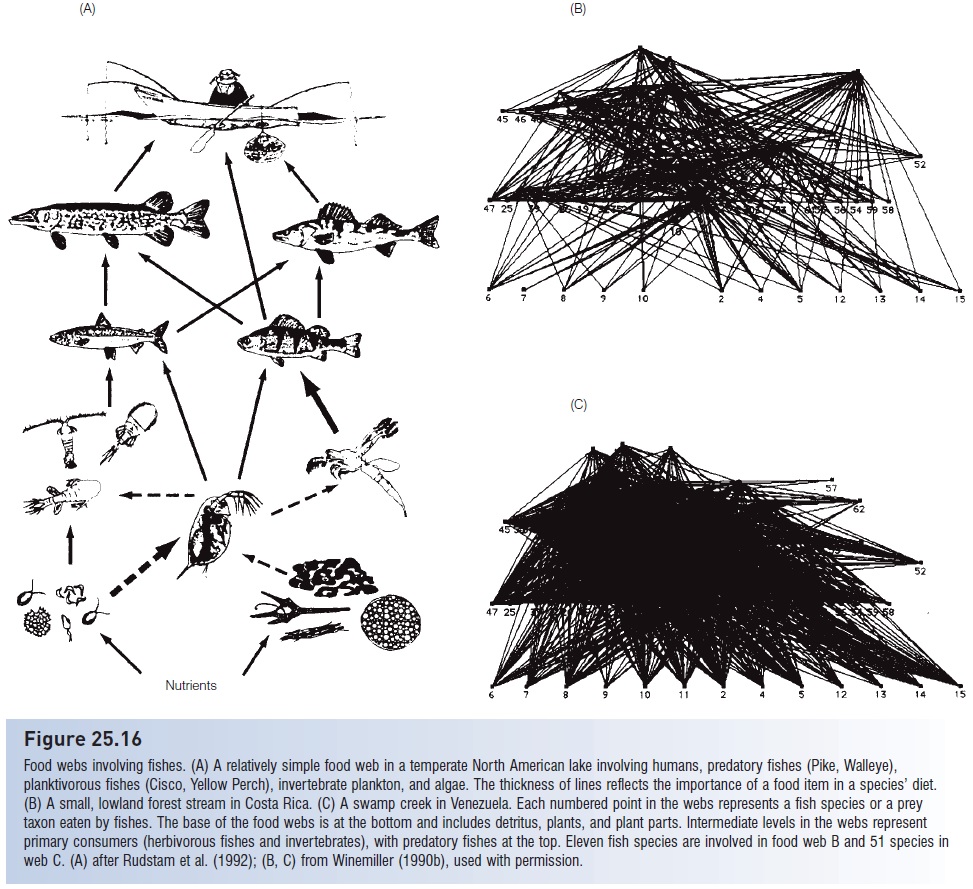
Figure 25.16
Food webs involving fishes. (A) A relatively simple food web in a temperate North American lake involving humans, predatory fishes (Pike, Walleye), planktivorous fishes (Cisco, Yellow Perch), invertebrate plankton, and algae. The thickness of lines reflects the importance of a food item in a species’ diet. (B) A small, lowland forest stream in Costa Rica. (C) A swamp creek in Venezuela. Each numbered point in the webs represents a fish species or a prey taxon eaten by fishes. The base of the food webs is at the bottom and includes detritus, plants, and plant parts. Intermediate levels in the webs represent primary consumers (herbivorous fishes and invertebrates), with predatory fishes at the top. Eleven fish species are involved in food web B and 51 species in web C. (A) after Rudstam et al. (1992); (B, C) from Winemiller (1990b), used with permission.
An often overlooked component of food webs concerns the placement of parasites, which are seldom included. The inclusion of parasites usually increases species richness, number of links, and food chain length plus two additional measures, connectance and nestedness, all which influence ecosystem stability (Lafferty et al. 2006). The web of feeding interactions in any community is undoubtedly tangled.
Most descriptions of trophic relationships within a community tend to characterize species as either relative specialists or generalists, referring to whether a species feeds predominantly on one or a few food types as compared to a species that feeds on many food types or even at several trophic levels. Specialist–generalist characterizations are used in general community descriptions and have also been invoked to explain the relatively high diversity of fishes and other taxa in tropical as compared to temperate communities. Tropical species are thought to be relatively specialized. The relatively narrow niches of specialists make it theoretically possible to fit more species into the resource spectrum of a habitat.
Although usually an accurate description, characterizing a species as either a specialist or a generalist may only apply to its habits under the feeding regime that exists in that habitat at the time of the description. An excellent example of this comes from a study of the feeding habits of two Panamanian toadfishes (Batrachoididae). Both species fed almost entirely (85–100%) on long-spined sea urchins, Diadema antillarum (Robertson 1987). However, Diadema underwent a massive die-off in early 1983 that wiped out 95–99% of the individuals across the Caribbean region. One would have predicted that such feeding specialists as the toadfishes would suffer population declines because of their relatively invariant feeding habits. However, populations of both species changed little, reproduction continued, and food habits shifted to a variety of mobile benthic invertebrates in one species and to fishes and mobile invertebrates in the other species. Hence these classic trophic specialists became relative generalists, sounding a cautionary note for anyone attempting to characterize community feeding relationships.
Related Topics