Chapter: Pharmaceutical Biotechnology: Fundamentals and Applications : Vaccines
Vaccine Design in Relation with the Immune Response
Vaccine
Design in Relation with theImmune Response
For the rational design of a new vaccine, under-standing of the
mechanisms of the protective immunity to the pathogen against which the
vaccine is developed is crucial. For instance, to prevent tetanus a high blood
titer of antibody against tetanus toxin is required; in mycobacterial diseases
such as tubercu-losis a macrophage-activating CMI is most effective; in case of
an influenza virus infection CTLs probably play a significant role.
Importantly, the immune effector mechanisms triggered by a vaccine and, hence,
the success of immunization not only depend on the nature of the protective
components but also on their presentation form, the presence of adjuvants, and
the route of administration.
The presentation form of the vaccine is one of the determinants that
influence the extent and type of immune response that will be evoked (Pashine
et al., 2005; Pulendran and Ahmed, 2006). DCs and other APCs play a pivotal
role in how the antigenic determinants of a vaccine will be processed and
presented to T-cells in the peripheral lymphoid organs. Through various PRRs,
DCs are more or less able to “sense” the type of pathogen that is encoun-tered.
This determines the set of co-stimulatorysignals and pro-inflammatory cytokines
that will be generated by APCs when presenting the antigen to Th-cells in the
peripheral lymphoid organs. For instance, pathogens or vaccines containing
lipopro-teins or peptidoglycans will trigger DCs via TLR-2, which predominantly
generates a Th2 response, whereas stimulation of DCs through TLR-3, -4, -5 or -8 is
known to yield robust Th1 responses. Therefore, vaccines should be formulated in such a way that
the appropriate Th response will be triggered. This can be done by presenting the antigen
in its native format, as is the case for the conventional vaccines, or by
adding adjuvants that stimulate the desired response (see below).
The response by B-cells is dependent upon the nature of the antigen and
two types of antigens can be distinguished:
1.
Thymus-independent antigens
include certain linear antigens that are not readily degraded in the body and
have a repeating determinant, such as bacterial polysaccharides. They are able
to stimulate B-cells without the Th-cell involvement. Thymus-independent antigens do not induce
immunological memory.
2.
Thymus-dependent antigens provoke
little or no antibody response in animals with few T-cells. Proteins are the
typical representatives of thymus-dependent antigens. A prerequisite for
thymus-dependency is that a physical linkage exists between the sites
recognized by B-cells and those by Th-cells. When a thymus-independent antigen is coupled to a carrier
protein containing Th-epitopes, it becomes thymus-dependent. As a result, these conjugates
are able to induce memory.
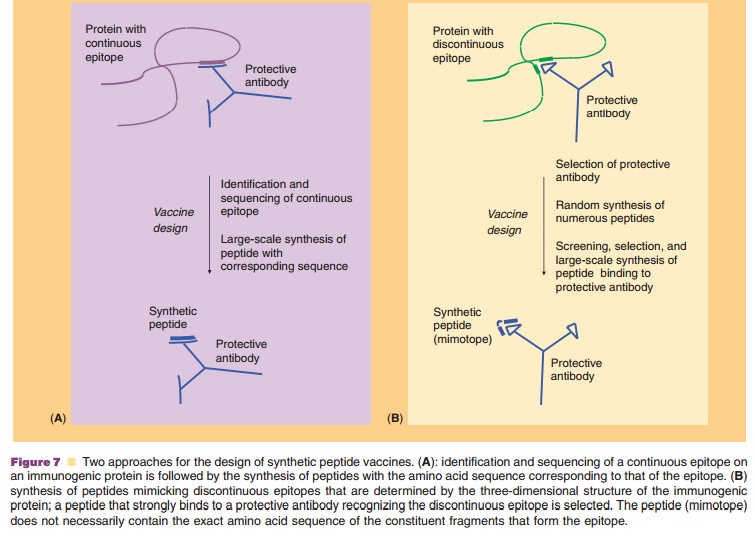
When the antigen is a protein, the epitopes can be continuous or discontinuous. Continuous epitopes involve linear peptide sequences (usually consisting of up to ten amino acid residues) of the protein (Fig. 7A). Discontinuous epitopes comprise amino acid residues sometimes far apart in the primary sequence, which are brought together through the unique folding of the protein (Fig. 7B). Antibody recognition of B-cell epitopes, whether continuous or discontinuous, is usually dependent on the conforma-tion (¼ three-dimensional structure). T-cell epitopes, on the other hand, are continuous peptide sequences, the conformation of which does not seem to play a role in T-cell recognition.
Related Topics