Chapter: 12th Microbiology : Chapter 12 : Microbial Genetics
Transfer of Genetic Material
Transfer of Genetic Material
Normally,
genes and the characteristics they code for are passed down from parent to
progeny. This is called vertical gene transfer. Bacteria and some lower
eukaryotes are unique in that they can pass DNA from one cell of the same
generation to another. The exchange of genes between two cells of the same
generation is referred to as horizontal gene transfer. Mechanisms like
transformation, transduction and conjugation takes place naturally and may
bring about genetic variation and genetic recombination. These gene transfer
mechanisms are also employed in genetic engineering to introduce desired gene
into the cells. Introducing a foreign gene or recombinant DNA into the cells is
one of the techniques used in genetic engineering. The success of cloning
depends on the efficiency of gene transfer process. The most commonly employed
gene transfer methods are transformation, conjugation, transduction,
electroporation, lipofection and direct transfer of DNA. The choice of the
method depends on the type of host cell (bacteria, fungi, plant, animal).
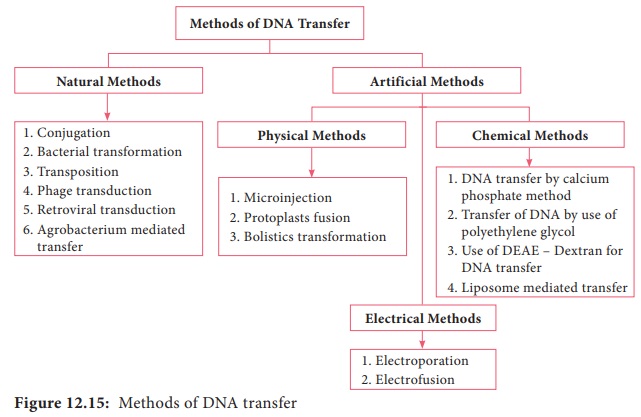
Figure 12.15 shows methods of DNA transfer
Transformation
Artificial competence can be
induced in laboratory by procedures
that involve making the cell passively permeable to DNA. Typically, the cells
are incubated in a solution containing divalent cations; most commonly, calcium
chloride solution under cold condition, which is then exposed toTransformation
is genetic alteration of a cell resulting from the direct uptake, incorporation
and expression of exogenous genetic material (exogenous DNA) from its
surroundings. Transformation occurs naturally in some species of bacteria, but
it can also take place by artificial means in other cells. For transformation
to happen, bacteria must be in a state of competence. Competence refers to the
state of being able to take up exogenous DNA from the environment. There are
two forms of competence: natural and artificial. Transformation works best with
DNA from closely- related species. The naturally-competent bacteria carry sets
of genes that provide the protein machinery to bring DNA across the cell
membrane(s).
There are
some differences in the mechanisms of DNA uptake by gram positive and gram
negative cells. However, they share some common features that involve related
proteins. The DNA first binds to the surface of the competent cells on a DNA
receptor, and passes through the cytoplasmic membrane via DNA translocase. Only
single stranded DNA may pass through, one strand is therefore degraded by
nucleases in the process, and the translocated single-stranded DNA may then be
integrated into the bacterial chromosomes. Figure 12.16 shows mechanism of
transformation
Artificial competence can be induced in laboratory by procedures that
involve making the cell passively permeable to DNA. Typically, the cells are
incubated in a solution containing divalent cations; most commonly, calcium
chloride solution under cold condition, which is then exposed to a pulse
of heat shock. Electroporation is another method of promoting competence. Using
this method, the cells are briefly shocked with an electric field of 10–20 kV/
cm which is thought to create holes in the cell membrane through which the
plasmid DNA may enter. After the electric shock, the holes are rapidly closed
by the cell’s membrane-repair mechanisms.
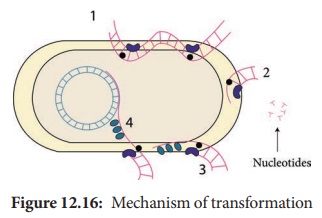
Figure 12.16: Mechanism of transformation
1. Binding of DNA; 2. Degradation of one strand; 3. Entry of ssDNA; 4. Integration into host chromosome.
Conjugation
The
initial evidence for bacterial conjugation, came from an experiment performed
by Joshua Lederberg and Edward L Tatum in 1946. Later in 1950, Bernard Davis
gave evidence that physical contact of the cells was necessary for conjugation.
During conjugation, two live bacteria (a donor and a recipient) come together,
join by cytoplasmic bridges (e.g. pilus) and transfer single stranded DNA (from
donor to recipient).
Inside
the recipient cell, the new DNA may integrate with the chromosome (rather rare)
or may remain free (as is the case with plasmids). Conjugation can occur among
the cells from different genera of bacteria, while transformation takes place
among the cells of a bacterial genus.
A plasmid called the fertility or F factor
plays a major role in conjugation. The F factor is about 100 kilobases long and
bears genes responsible for cell attachment and plasmid transfer between
specific bacterial strains during conjugation. F factor is made up of
a. tra region (tra operon / transfer genes): genes
coding the F pilus and DNA transfer,
b. Insertion sequence: genes assisting plasmid
integration into host cell chromosome.
Thus, the F factor is
an episome -
a genetic material that can exist outside the bacterial chromosome or be
integrated into it.
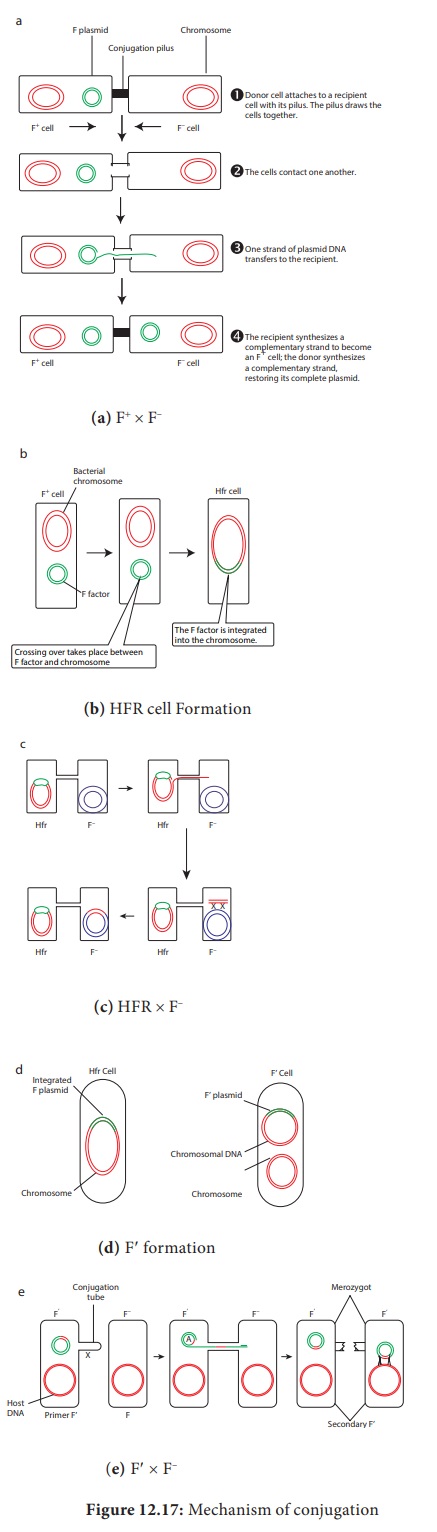
During F+ × F –
mating or conjugation (Figure 12.17a) the F factor replicates by the rolling
circle mechanism and a copy moves to the recipient. The channel for DNA
transfer could be either the hollow F pilus or a special conjugation bridge
formed upon contact. The entering strand is copied to produce double – stranded
DNA.
F factor can integrate into the bacterial
chromosome at several different locations by recombination between homologous
insertion sequences present on both the plasmid and host chromosomes. The
integration of F factor into bacterial chromosome results in formation of HFR
(High Frequency Recombination) cell. When integrated, the Fplasmid’s tra operon
is still functional; the plasmid can direct the synthesis of pili, carry out
rolling circle replication, and transfer genetic material to an F- recipient
cell. An HFR cell is so called because it exhibits a very high efficiency of
chromosomal gene transfer in comparison with F+ cells. In F+ cells the independent F factor rarely transfer
chromosomal genes hence the recombination frequency is low. Figure 12.17b shows
formation of HFR cell. When an HFR cell is mated with F- cell the F- recipient
does not become F+ unless
the whole chromosome is transferred as explained in Figure 12.17c. The
connection usually breaks before this process is finished. Thus, complete F
factor usually is not transferred, and the recipient remains F-.
Because
the F plasmid is an episome, it can leave (deintegrate) the bacterial
chromosome. Sometimes during this process, the plasmid makes an error in
excision and picks up a portion of the chromosomal material to form an F′ plasmid.
Figure 12.17d shows formation of F′. During F′XF- conjugation (Figure 12.17e) the recipient becomes
F′ and is a
partially diploid since it has two set of the genes carried by the plasmid.
The natural phenomenon of conjugation is now
exploited for gene transfer and Recombinant DNA technology. In general, the
plasmids lack conjugative functions and therefore, they are not as such capable
of transferring DNA to the recipient cells. However, some plasmids with
conjugative properties can be prepared and used.
Transduction
Transduction
is the transfer of bacterial genes from one bacteria to other by viruses.
Example: Bacteriophage (Bacterial viruses). To understand the role of
bacteriophage in gene transfer, the lifecycle of bacteriophage is described
below briefly.
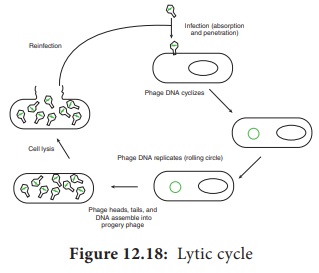
After
infecting the host cell, a bacteriophage (phage for short) often takes control
and forces the host to make many copies of the virus. Eventually the host
bacterium bursts or lyses and releases new phages. This reproductive cycle is
called a lytic cycle because it ends in lysis of the host.
The lytic
cycle (Figure 12.18) has four phases.
1. Attachment - Virus particle attaches to a
specific receptor site on the bacterial surface.
2. Penetration - the genetic material, which is
often double stranded DNA, then enters the cell.
3. Biosynthesis - After adsorption and penetration,
the virus chromosome forces the bacterium to make viral components-viral
nucleic acids and proteins.
4. Assembly - Phages are assembled from the virus
components. Phage nucleic acid is packed within the virus’s protein coat.
5. Release - mature viruses are released by cell lysis.
Bacterial
viruses that reproduce using a lytic cycle often are called virulent bacteriophages (e.g. T phages) because they destroy the host cell. The genome of many DNA phages such as
the lambda phage, after adsorption and penetration do not take control of its
host and does not destroy the host. Instead the viral genome remains within the
host cell and is reproduced along with the bacterial chromosome. The infected
bacteria may multiply for long periods while appearing perfectly normal. Each
of these infected bacteria can produce phages and lyses under appropriate
environmental conditions. This relationship between phage and its host is
called lysogeny (Figure 12.19).
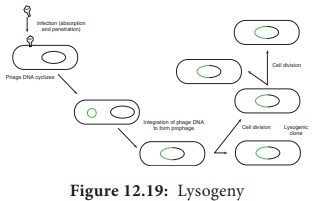
Bactera that
can produce phage particles under some conditions are said to be lysogens or lysogenic bacteria. Phages which
are able to establish lysogeny are called temperate
phages
The
latent form of virus genome that remains within the host without destroying the
host is called the prophage.
The
prophage usually is integrated into the bacterial genome. Sometimes phage
reproduction is triggered in a lysogenized culture by exposure to UV radiation
or other factors. The lysosomes are then destroyed and new phages released –
This phenomenon is called induction (Figure 12.20).
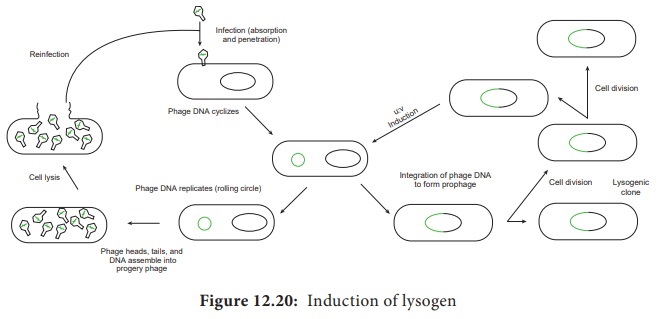
Sometimes,
bacterial genes are incorporated into a phage capsid because of errors made
during the virus life cycle. The virus containing these genes then infects them
into another bacterium, resulting in the transfer of genes from one bacterium
to the other. Transduction may be the most common mechanism for gene exchange
and recombination in bacteria.
There are two very different kinds of
transduction.
1. Generalized transduction
2. Specialized
transduction
Generalized transduction
(Figure
12.21a) occurs during the lytic cycle of virulent and temperate phages. During
the assembly stage, when the viral chromosomes are packaged into protein
capsids, random fragments of the partially degraded bacterial chromosome also
may be packaged by mistake. The resulting virus particles often injects the DNA
into another bacterial cell but does not initiate a lytic cycle. Thus in
generalized transduction any part of the bacterial chromosome can be
transferred. Once the DNA has been injected it may integrate into the recipient
cell’s chromosome to preserve the transferred genes. About 70 to 90% of the
transferred DNA is not integrated but is often able to survive and express
itself. However, if the transferred DNA is degraded gene transfer is
unsuccessful.
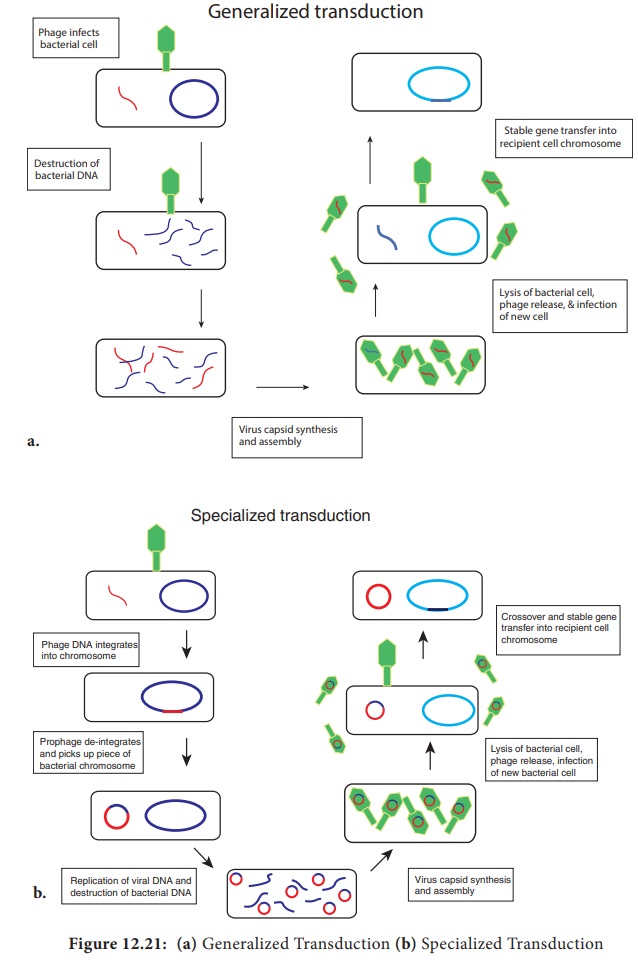
Specialized Transduction (Figure 12.21b) is also called restricted transduction in which only specific portions of the bacterial genome is carried by the phage. When a prophage is induced to leave the host chromosome, exicision is sometimes carried out improperly. The resulting phage genome contains portions of the bacterial chromosome next to the integration site. When this phage infects another bacterium, it transfers the bacterial genes from the donor bacterium along with phage DNA. Here only the bacterial genes that are close to the site of prophage are transferred. So, this transduction is called specialized.
Infobits
The chromosomes of bacteria, viruses, and eukaryotic cells
contain pieces of DNA that move around the genome. Such movement is called
transposition. DNA segments that carry the genes required for this process and
consequently move about chromosomes are transposable elements or transposons.
Transposons are also called jumping genes because they can jump from one DNA to
another, resulting in mutation of the cell. They were first discovered in 1951
by Barbara McClintock whose significant discovery was ignored by scientific
community for many years. She was awarded the Nobel Prize in 1983.
Related Topics