Chapter: Plant Biochemistry: Phloem transport distributes photoassimilates to the various sites of consumption and storage
There are two modes of phloem loading
There are two modes of phloem loading
Photoassimilates generated in the mesophyll cells, such as sucrose, various oligosaccharides, polyols as well as amino acids, diffuse via plasmodesmata to the bundle sheath cells. The subsequent transport of photoassimilates from the bundle sheath cells to the sieve tubes can occur in two different ways:
1. Plants in which oligosaccharides from the raffinose family are translocated into the sieve tubes (e.g., squash plants), the bundle sheath cells are connected to specialized companion cells (intermediary cells) and further to the sieve tubes via a large number of plasmodes-mata. These plants transfer the photoassimilates to the sieve tubes via plasmodesmata in a process termed symplastic phloem loading.
2. In contrast, in apoplastic phloem loading, found for instance in the leaves of cereals, sugar beet, rapeseed, and potato, photoassimilates are first transported from the source cells via the bundle sheath cells to the extracellular compartment, the apoplast, and then by active transport into the sieve tube compartment (Fig. 13.3). Since the concentration of sucrose, polyols and amino acids in the source cells is very much higher than in the apoplast, this export does not seem to require any energy input. The translocators mediating the export from the bundle sheath to the apoplast have not yet been characterized.
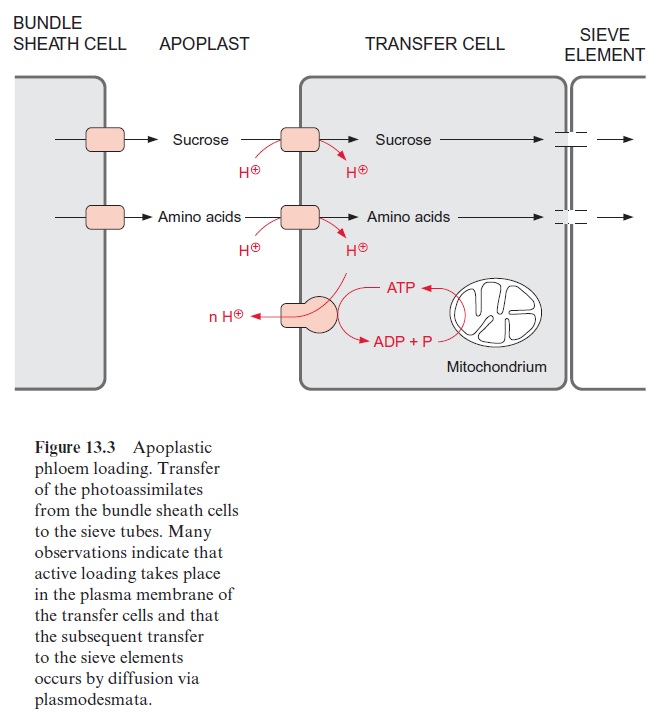
The companion cells participating in the apoplastic phloem loading are termed transfer cells. The transport of sucrose and amino acids from the apo-plasts to the phloem proceeds via a proton symport (Fig. 13.3). This is driven by a proton gradient between the apoplast and the interior of the companion cells and the sieve tubes. The proton gradient is generated by an H+ -P ATPase present in the plasma membrane. The required ATP is pro-duced by mitochondrial oxidation. By now a number of H+ -driven symport-ers involved in the phloem loading of sucrose, polyols and amino acids have been identified and characterized in several plants. In the vascular bundles of Plantago major using specific antibodies, an H+ -sucrose translocator and two H+ -polyol translocators have been localized in the plasma membrane of trans-fer cells (Fig. 13.3). The substrates for mitochondrial respiration are provided by degradation of sucrose via sucrose synthase (see also Fig. 13.5) to hexose phosphates, which are further degraded by glycolytic metabolism. Another substrate for respiration is glutamate . In many plants this amino acid is present in relatively high concentrations in the phloem sap.
In the plants with apoplastic phloem loading, sucrose, also in combina-tions with the polyols sorbitol and mannitol, is the transport form for car-bohydrates (hexoses are not transported), while no special transport form exists for amino nitrogen. In principle, all protein-building amino acids are transported. The ratio of a single amino acid versus the sum of amino acids is very similar in the phloem sap and in the source cells. The amino acids most frequently found in the phloem sap are glutamate, glutamine, and aspartate, but also much alanine is found in some plants. Several amino acid translocators with a broad specificity for various amino acids were identified, which are presumed to participate in the phloem loading.
Related Topics