Chapter: Medical Physiology: Regulation of Acid-Base Balance
Secretion of Hydrogen Ions and Reabsorption of Bicarbonate Ions by the Renal Tubules
Secretion of Hydrogen Ions and Reabsorption of Bicarbonate Ions by the Renal Tubules
Hydrogen ion secretion and bicarbonate reabsorption occur in virtually all parts of the tubules except the descending and ascending thin limbs of the loop of Henle. Figure 30–4 summarizes bicarbonate reabsorp-tion along the tubule. Keep in mind that for each bicar-bonate reabsorbed, an H+ must be secreted.
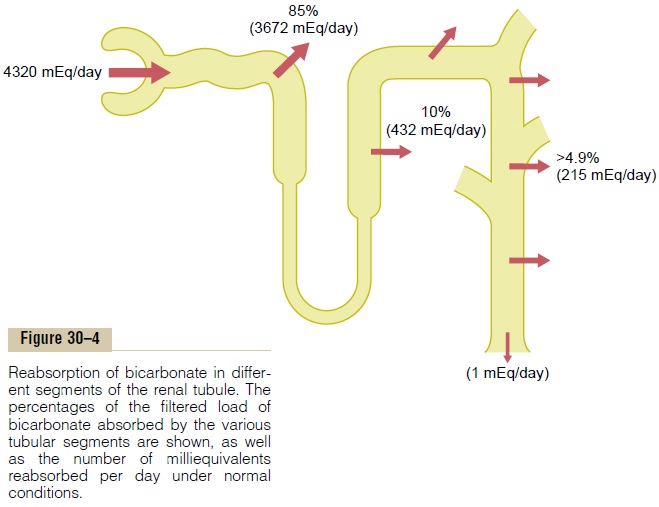
About 80 to 90 per cent of the bicarbonate reab-sorption (and H+ secretion) occurs in the proximal tubule, so that only a small amount of bicarbonate flows into the distal tubules and collecting ducts. In the thick ascending loop of Henle, another 10 per cent of the filtered bicarbonate is reabsorbed, and the remainder of the reabsorption takes place in the distal tubule and collecting duct.
As discussed previously, the mech-anism by which bicarbonate is reabsorbed also involves tubular secretion of H+, but different tubular segments accomplish this task differently.
Hydrogen Ions Are Secreted by Secondary Active Transport in the Early Tubular Segments
The epithelial cells of the proximal tubule, the thick segment of the ascending loop of Henle, and the early distal tubule all secrete H+ into the tubular fluid by sodium-hydrogen counter-transport, as shown in Figure 30–5. This secondary active secretion of H+ is coupled with the transport of Na+ into the cell at the luminal membrane by the sodium-hydrogen exchanger protein, and the energy for H+ secretion against a concentration gradient is derived from the sodium gradient favoring Na+ movement into the cell. This gradient is established by the sodium-potassium adenosine triphosphatase (ATPase) pump in the baso-lateral membrane. More than 90 per cent of the bicar-bonate is reabsorbed in this manner, requiring about 3900 milliequivalents of H+ to be secreted each day by the tubules. This mechanism, however, does not estab-lish a very high H+ concentration in the tubular fluid; the tubular fluid becomes very acidic only in the col-lecting tubules and collecting ducts.
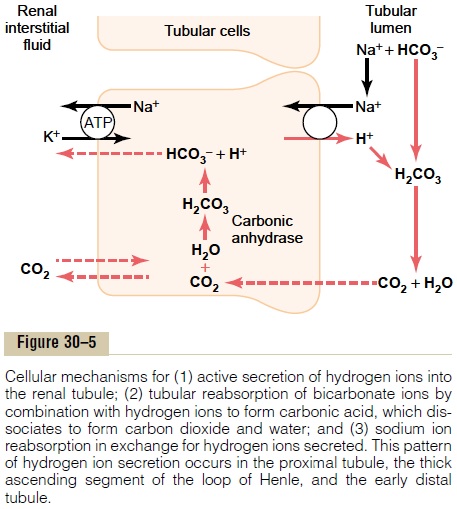
Figure 30–5 shows how the process of H+ secretion achieves bicarbonate reabsorption. The secretory process begins when CO2 either diffuses into the tubular cells or is formed by metabolism in the tubular epithelial cells. CO2, under the influence of the enzyme carbonic anhydrase, combines with H2O to formH2CO3, which dissociates into HCO3– and H+. The H+ is secreted from the cell into the tubular lumen by sodium-hydrogen counter-transport. That is, when an Na+ moves from the lumen of the tubule to the inte-rior of the cell, it first combines with a carrier protein in the luminal border of the cell membrane; at the same time, an H+ in the interior of the cells combines with the carrier protein. The Na+moves into the cell down a concentration gradient that has been estab-lished by the sodium-potassium ATPase pump in the basolateral membrane. The gradient for Na+ move-ment into the cell then provides the energy for moving H+ in the opposite direction from the interior of the cell to the tubular lumen.
The HCO3– generated in the cell (when H+ dissoci-ates from H2CO3) then moves downhill across the basolateral membrane into the renal interstitial fluid and the peritubular capillary blood. The net result is that for every H+ secreted into the tubular lumen, an HCO3– enters the blood.
Filtered Bicarbonate Ions Are Reabsorbed by Interaction with Hydrogen Ions in the Tubules
Bicarbonate ions do not readily permeate the luminal membranes of the renal tubular cells; therefore, HCO3– that is filtered by the glomerulus cannot be directly reabsorbed. Instead, HCO3– is reabsorbed by a special process in which it first combines with H+ to form H2CO3, which eventually becomes CO2 and H2O, as shown in Figure 30–5.
This reabsorption of HCO3– is initiated by a reaction in the tubules between HCO3– filtered at the glomeru-lus and H+ secreted by the tubular cells. The H2CO3 formed then dissociates into CO2 and H2O. The CO2 can move easily across the tubular membrane; there-fore, it instantly diffuses into the tubular cell, where it recombines with H2O, under the influence of carbonic anhydrase, to generate a new H2CO3 molecule. This H2CO3 in turn dissociates to form HCO3– and H+; the HCO3– then diffuses through the basolateral mem-brane into the interstitial fluid and is taken up into the peritubular capillary blood. The transport of HCO3 across the basolateral membrane is facilitated by two mechanisms: (1) Na+-HCO3– co-transport and (2) Cl–-HCO3– exchange.
Thus, each time an H+ is formed in the tubular epithe-lial cells, an HCO3- is also formed and released back into the blood. The net effect of these reactions is“reabsorption” of HCO3– from the tubules, although the HCO3– that actually enters the extracellular fluid is not the same as that filtered into the tubules. The reabsorption of filtered HCO3– does not result in net secretion of H+ because the secreted H+ combines with the filtered HCO3– and is therefore not excreted.
Bicarbonate Ions Are “Titrated” Against Hydrogen Ions in the Tubules. Under normal conditions, the rate of tubularH+ secretion is about 4400 mEq/day, and the rate of fil-tration by HCO3– is about 4320 mEq/day. Thus, the quantities of these two ions entering the tubules are almost equal, and they combine with each other to form CO2 and H2O. Therefore, it is said that HCO3– and H+ normally “titrate” each other in the tubules.
The titration process is not quite exact because there is usually a slight excess of H+ in the tubules to be excreted in the urine. This excess H+ (about 80 mEq/ day) rids the body of nonvolatile acids produced by metabolism. As discussed later, most of this H+ is not excreted as free H+ but rather in combination with other urinary buffers, especially phosphate and ammonia.
When there is an excess of HCO3– over H+ in the urine, as occurs in metabolic alkalosis, the excess HCO3– cannot be reabsorbed; therefore, the excess HCO3– is left in the tubules and eventually excreted into the urine, which helps correct the metabolic alkalosis.
In acidosis, there is excess H+ relative to HCO3–, causing complete reabsorption of the bicarbonate; the excess H+ passes into the urine. The excess H+ is buffered in the tubules by phosphate and ammonia and eventually excreted as salts. Thus, the basic mech-anism by which the kidneys correct either acidosis or alkalosis is incomplete titration of H+ against HCO3–, leaving one or the other to pass into the urine and be removed from the extracellular fluid.
Primary Active Secretion of Hydrogen Ions in the Intercalated Cells of Late Distal and Collecting Tubules
Beginning in the late distal tubules and continuing through the remainder of the tubular system, the tubular epithelium secretes H+ by primary activetransport. The characteristics of this transport are dif-ferent from those discussed for the proximal tubule, loop of Henle, and early distal tubule.
The mechanism for primary active H+ secretion is shown in Figure 30–6. It occurs at the luminal mem-brane of the tubular cell, where H+ is transported directly by a specific protein, a hydrogen-transporting ATPase. The energy required for pumping the H+ is derived from the breakdown of ATP to adenosine diphosphate.
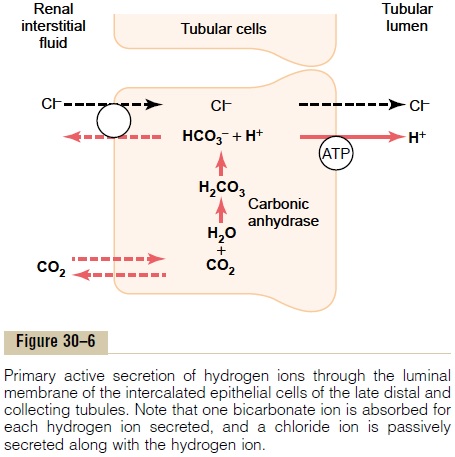
Primary active secretion of H+ occurs in a special type of cell called the intercalated cells of the late distal tubule and in the collecting tubules. Hydrogen ion secretion in these cells is accomplished in two steps: (1) the dissolved CO2 in this cell combines with H2O to form H2CO3, and (2) the H2CO3 then dissociates into HCO3–, which is reabsorbed into the blood, plus H+, which is secreted into the tubule by means of the hydrogen-ATPase mechanism. For each H+ secreted, an HCO3– is reabsorbed, similar to the process in the proximal tubules. The main difference is that H+ moves across the luminal membrane by an active H+ pump
Although the secretion of H+ in the late distal tubule and collecting tubules accounts for only about 5 per cent of the total H+ secreted, this mechanism is impor-tant in forming a maximally acidic urine. In the prox-imal tubules, H+ concentration can be increased only about threefold to fourfold, and the tubular fluid pH can be reduced to only about 6.7, although large amounts of H+are secreted by this nephron segment.However, H+ concentration can be increased as much as 900-fold in the collecting tubules. This decreases the pH of the tubular fluid to about 4.5, which is the lower limit of pH that can be achieved in normal kidneys.
Related Topics