Chapter: Medical Physiology: Cortical and Brain Stem Control of Motor Function
Role of the Brain Stem in Controlling Motor Function
Role of the Brain Stem in Controlling Motor Function
The brain stem consists of the medulla, pons, and mes-encephalon. In one sense, it is an extension of thespinal cord upward into the cranial cavity, because it contains motor and sensory nuclei that perform motor and sensory functions for the face and head regions in the same way that the spinal cord performs these func-tions from the neck down. But in another sense, the brain stem is its own master because it provides many special control functions, such as the following:
1. Control of respiration
2. Control of the cardiovascular system
3. Partial control of gastrointestinal function
4. Control of many stereotyped movements of the body
5. Control of equilibrium
6. Control of eye movements
Finally, the brain stem serves as a way station for “command signals” from higher neural centers. In the following sections, we discuss the role of the brain stem in controlling whole-body movement and equilibrium. Especially important for these purposes are the brain stem’s reticular nuclei and vestibular nuclei.
Support of the Body Against Gravity— Roles of the Reticular and Vestibular Nuclei
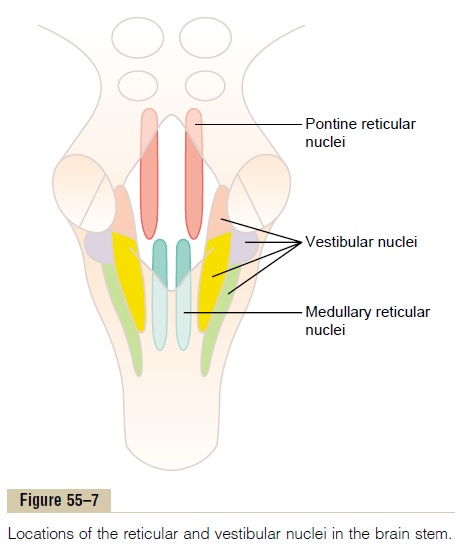
Figure 55–7 shows the locations of the reticular and vestibular nuclei in the brain stem.
Excitatory-Inhibitory Antagonism Between Pontine and Medullary Reticular Nuclei
The reticular nuclei are divided into two major groups:
(1) pontine reticular nuclei, located slightly posteriorly and laterally in the pons and extending into the mes-encephalon, and (2) medullary reticular nuclei, which extend through the entire medulla, lying ventrally and medially near the midline. These two sets of nuclei function mainly antagonistically to each other, with the pontine exciting the antigravity muscles and the medullary relaxing these same muscles.
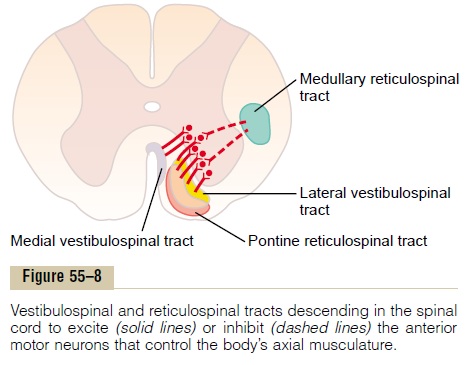
Pontine Reticular System. The pontine reticular nucleitransmit excitatory signals downward into the cord through the pontine reticulospinal tract in the anterior column of the cord, as shown in Figure 55–8. The fibers of this pathway terminate on the medial anterior motor neurons that excite the axial muscles of the body, which support the body against gravity—that is, the muscles of the vertebral column and the extensor muscles of the limbs.
The pontine reticular nuclei have a high degree of natural excitability. In addition, they receive strong excitatory signals from the vestibular nuclei, as well as from deep nuclei of the cerebellum. Therefore, when the pontine reticular excitatory system is unopposed by the medullary reticular system, it causes powerful excitation of antigravity muscles throughout the body, so much so that four-legged animals can be placed in a standing position, supporting the body against gravity without any signals from higher levels of the brain.
Medullary Reticular System. The medullary reticularnuclei transmit inhibitory signals to the same anti-gravity anterior motor neurons by way of a different tract, the medullary reticulospinal tract, located in the lateral column of the cord, as also shown in Figure 55–8. The medullary reticular nuclei receive strong input collaterals from (1) the corticospinal tract, (2) the rubrospinal tract, and (3) other motor pathways. These normally activate the medullary reticular inhibitory system to counterbalance the excitatory signals from the pontine reticular system, so that under normal conditions, the body muscles are not abnor-mally tense.
Yet some signals from higher areas of the brain can “disinhibit” the medullary system when the brain wishes to excite the pontine system to cause standing. At other times, excitation of the medullary reticular system can inhibit antigravity muscles in certain por-tions of the body to allow those portions to perform special motor activities. The excitatory and inhibitory reticular nuclei constitute a controllable system that is manipulated by motor signals from the cerebral cortex and elsewhere to provide necessary background muscle contractions for standing against gravity and to inhibit appropriate groups of muscles as needed so that other functions can be performed.
Role of the Vestibular Nuclei to Excite the Antigravity Muscles All the vestibular nuclei, shown in Figure 55–7, func-tion in association with the pontine reticular nuclei to control the antigravity muscles. The vestibular nuclei transmit strong excitatory signals to the antigravity muscles by way of the lateral and medial vestibu-lospinal tracts in the anterior columns of the spinalcord, as shown in Figure 55–8. Without this support of the vestibular nuclei, the pontine reticular system would lose much of its excitation of the axial anti-gravity muscles.
The specific role of the vestibular nuclei, however, is to selectively control the excitatory signals to the dif-ferent antigravity muscles to maintain equilibrium inresponse to signals from the vestibular apparatus.
The Decerebrate Animal Develops Spastic Rigidity
When the brain stem of an animal is sectioned below the midlevel of the mesencephalon, but the pontine and medullary reticular systems as well as the vestibular system are left intact, the animal develops a condition called decerebrate rigidity. This rigidity does not occur in all muscles of the body but does occur in the anti-gravity muscles—the muscles of the neck and trunk and the extensors of the legs.
The cause of decerebrate rigidity is blockage of nor-mally strong input to the medullary reticular nuclei from the cerebral cortex, the red nuclei, and the basal ganglia. Lacking this input, the medullary reticular inhibitor system becomes nonfunctional; full overactiv-ity of the pontine excitatory system occurs, and rigidity develops. We shall see later that other causes of rigidity occur in other neuromotor diseases, especially lesions of the basal ganglia.
Related Topics