Chapter: Medical Physiology: Motor Functions of the Spinal Cord; the Cord Reflexes
Receptor Function of the Muscle Spindle
Receptor Function of the Muscle Spindle
Structure and Motor Innervation of the Muscle Spindle. The organization of the muscle spindle is shown in Figure 54–2. Each spindle is 3 to 10 millimeters long. It is built around 3 to 12 very small intrafusal muscle fibers that are pointed at their ends and attached to the glycoca-lyx of the surrounding large extrafusal skeletal muscle fibers.
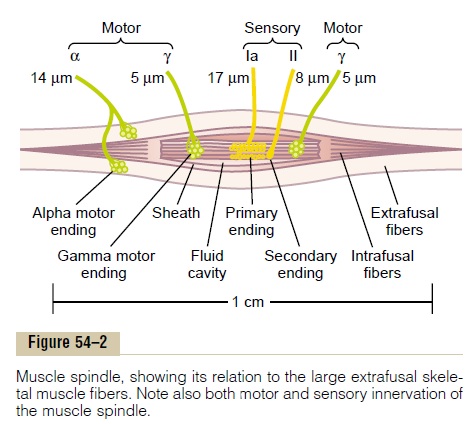
Each intrafusal muscle fiber is a very small skeletal muscle fiber. However, the central region of each of these fibers—that is, the area midway between its two ends—has few or no actin and myosin fila-ments.Therefore, this central portion does not contract when the ends do. Instead, it functions as a sensory receptor, as described later. The end portions that do contract are excited by small gamma motor nervefibers that originate from small type A gamma motorneurons in the anterior horns of the spinal cord, as described earlier. These gamma motor nerve fibers are also called gamma efferent fibers, in contradistinc-tion to the large alpha efferent fibers (type A alpha nerve fibers) that innervate the extrafusal skeletal muscle.
Sensory Innervation of the Muscle Spindle. The receptorportion of the muscle spindle is its central portion. In this area, the intrafusal muscle fibers do not have myosin and actin contractile elements. As shown in Figure 54–2 and in more detail in Figure 54–3, sensory fibers originate in this area. They are stimulated by stretching of this midportion of the spindle. One can readily see that the muscle spindle receptor can be excited in two ways:
1. Lengthening the whole muscle stretches the midportion of the spindle and, therefore, excites the receptor.
2. Even if the length of the entire muscle does not change, contraction of the end portions of the spindle’s intrafusal fibers stretches the midportion of the spindle and therefore excites the receptor.
Two types of sensory endings are found in this central receptor area of the muscle spindle. They are the primary ending and thesecondary ending.
Primary Ending. In the center of the receptor area, alarge sensory nerve fiber encircles the central portion of each intrafusal fiber, forming the so-called primaryending or annulospiral ending. This nerve fiber is atype Ia fiber averaging 17 micrometers in diameter, and it transmits sensory signals to the spinal cord at a velocity of 70 to 120 m/sec, as rapidly as any type of nerve fiber in the entire body.
Secondary Ending. Usually one but sometimes twosmaller sensory nerve fibers—type II fibers with an average diameter of 8 micrometers—innervate the receptor region on one or both sides of the primary ending, as shown in Figures 54–2 and 54–3. This sensory ending is called the secondary ending; some-times it encircles the intrafusal fibers in the same way that the type Ia fiber does, but often it spreads like branches on a bush.
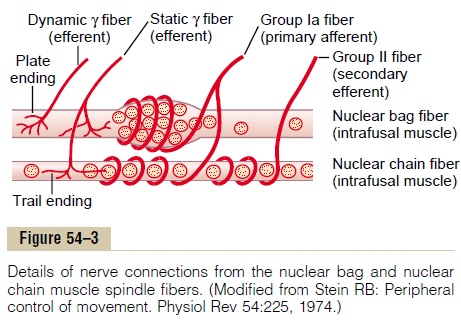
Division of the Intrafusal Fibers into Nuclear Bag and Nuclear Chain Fibers—Dynamic and Static Responses of the Muscle Spindle. There are also two types of muscle spindleintrafusal fibers: (1) nuclear bag muscle fibers (one to three in each spindle), in which several muscle fiber nuclei are congregated in expanded “bags” in the central portion of the receptor area, as shown by the top fiber in Figure 54–3, and (2) nuclear chain fibers (three to nine), which are about half as large in diam-eter and half as long as the nuclear bag fibers and have nuclei aligned in a chain throughout the receptor area, as shown by the bottom fiber in the figure. The primary sensory nerve ending (the 17-micrometer sensory fiber) is excited by both the nuclear bag intrafusal fibers and the nuclear chain fibers. Conversely, the sec-ondary ending (the 8-micrometer sensory fiber) is usually excited only by nuclear chain fibers.These rela-tions are shown in Figure 54–3.
Response of Both the Primary and the Secondary Endings to the Length of the Receptor—“Static” Response. When the receptor portion of the musclespindle is stretched slowly, the number of impulses transmitted from both the primary and the secondary endings increases almost directly in proportion to the degree of stretching, and the endings continue to trans-mit these impulses for several minutes. This effect is called the static response of the spindle receptor, meaning simply that both the primary and secondary endings continue to transmit their signals for at least several minutes if the muscle spindle itself remains stretched.
Response of the Primary Ending (but Not the Sec-ondary Ending) to Rate of Change of Receptor Length—“Dynamic” Response. When the length ofthe spindle receptor increases suddenly, the primary ending (but not the secondary ending) is stimulated especially powerfully. This excess stimulus of the primary ending is called the dynamic response, which means that the primary ending responds extremely actively to a rapid rate of change in spindle length. Even when the length of a spindle receptor increases only a fraction of a micrometer, if this increase occurs in a fraction of a second, the primary receptor trans-mits tremendous numbers of excess impulses to the large 17-micrometer sensory nerve fiber, but onlywhile the length is actually increasing. As soon as thelength stops increasing, this extra rate of impulse dis-charge returns to the level of the much smaller static response that is still present in the signal.
Conversely, when the spindle receptor shortens, exactly opposite sensory signals occur. Thus, the primary ending sends extremely strong, either positive or negative, signals to the spinal cord to apprise it of any change in length of the spindle receptor.
Control of Intensity of the Static and Dynamic Responses by the Gamma Motor Nerves. The gamma motor nerves to themuscle spindle can be divided into two types: gamma-dynamic (gamma-d) and gamma-static (gamma-s).The first of these excites mainly the nuclear bag intrafusal fibers, and the second excites mainly the nuclear chain intrafusal fibers. When the gamma-d fibers excite the nuclear bag fibers, the dynamic response of the muscle spindle becomes tremendously enhanced, whereas the static response is hardly affected. Conversely, stimula-tion of the gamma-s fibers, which excite the nuclear chain fibers, enhances the static response while having little influence on the dynamic response. Subsequent paragraphs illustrate that these two types of muscle spindle responses are important in different types of muscle control.
Continuous Discharge of the Muscle Spindles Under Normal Conditions. Normally, particularly when there is somedegree of gamma nerve excitation, the muscle spindles emit sensory nerve impulses continuously. Stretching the muscle spindles increases the rate of firing, whereas shortening the spindle decreases the rate of firing. Thus, the spindles can send to the spinal cord either positive signals—that is, increased numbers of impulses to indicate stretch of a muscle—or negativesignals—below-normal numbers of impulses to indi-cate that the muscle is unstretched.
Related Topics