Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Individuals, populations, and assemblages
Predation - Diversity of Fishes: Assemblages
Predation
Predator–prey interactions among species in an assemblage can have direct and indirect effects on prey population size and distribution. Direct effects include immediate mortal - ity or delayed mortality due to injury. Indirect effects involve habitat shifts caused by a predator’s presence that force potential prey to use suboptimal habitats, which can affect individual growth and reproduction (see above). Population-level responses associated with predation are usually density dependent and vary with the age of the prey. Density-dependent changes occur when the size of the prey population determines the impact of the predator. Direct density dependence is referred to as compensatory, meaning that predation increases to compensate for increases in prey population size. Predation by seabirds on schooling pelagic fishes is often compensatory in both the short and long term. The feeding activities of one bird draws the attention of other birds and the number of predators arriving at the site increases in direct relation to the size of the fish school on which they are feeding. Successful feeding by the birds in turn increases survivorship of their young, which means an increase in predators in the next generation, all dependent on the size of the fish resource.
Intercohort cannibalism, in which older fish eat younger age classes, can have a strong density-dependent impact on year class strength. Consider a population with three year classes. A large cannibalistic cohort can depress the numbers of the next, younger cohort. When the younger cohort reaches a size where it is a threat to the third, youngest age class, reduced numbers in the second age group have relatively little impact, which translates into high survivorship in the third group. In this way, population cycles can be established through the density-dependent effects of cannibalism. Just this type of scenario has been invoked to explain 2-year cycles of abundance in Pink Salmon, Oncorhynchus gorbuscha, in the Pacific Northwest (Ricker 1962).
Inverse density dependence is considered depensatory because relative predation risk and impact decrease as prey numbers increase. Depensatory predation occurs when a fixed number of predators become swamped or saturated by large numbers of prey. Under such conditions, the proportion of the prey captured decreases as prey numbers increase. For example, an individual salmon smolt that is migrating to the sea reduces its risk of death if it can time its downstream migration to coincide with that of other smolts, since predators take only a small number of migrants. By extension, the proportion of the prey population killed decreases as the population increases (Wootton 1990).
Regardless of the nature of the relationship between predator and prey densities, predation can have dramatic effects on prey population size. It is generally held that most of the mortality in eggs and larvae of species with planktonic young is due to predation, with predators taking >99% of the individuals (Bailey & Houde 1989; see above). When older, fish are still subject to predation, but the threat falls off progressively with increasing age and size, forming what is described as exponentially declining mortality (Fig. 24.8). Refuge availability may influence the impact that predators have on later life history stages. Observations and experimental manipulations on coral reefs indicate that prey population density is directly related to the number and availability of holes where prey can hide, which also implies that competition for refuge sites could interact with predation to determine population density and diversity. Several experimental studies on reef fishes have in fact shown that removing predators leads to increased density of prey, reinforcing descriptions of dramatic trophic cascades (see Indirect effects and trophic cascades). The effects of predator density on prey diversity may follow a similar pattern (Hixon 1991; Hixon & Beets 1993). Regardless, regulation of population size ultimately involves an interplay of competition and predation, often mediated by habitat availability (e.g., Holbrook & Schmitt 2002; Hixon & Jones 2005).
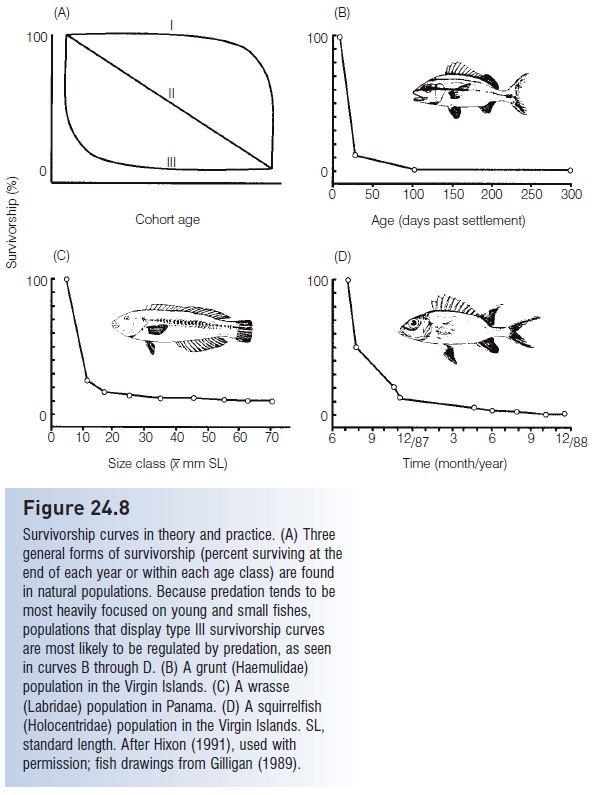
Figure 24.8
Survivorship curves in theory and practice. (A) Three general forms of survivorship (percent surviving at the end of each year or within each age class) are found in natural populations. Because predation tends to be most heavily focused on young and small fishes, populations that display type III survivorship curves are most likely to be regulated by predation, as seen in curves B through D. (B) A grunt (Haemulidae) population in the Virgin Islands. (C) A wrasse (Labridae) population in Panama. (D) A squirrelfish (Holocentridae) population in the Virgin Islands. SL, standard length. After Hixon (1991), used with permission; fish drawings from Gilligan (1989).
Predation can also affect gene frequencies in populations through the evolution of antipredator adaptations. Guppies
in small streams in Venezuela and on the island of Trinidad occur in pools that differ in levels of predation. Upstream areas tend to have few if any predators, often limited to a single topminnow species, Rivulus marmoratus (Cyprinodontidae). Further downstream, more predators occur, including a cichlid (Crenicichla), a characin (Hoplias), and freshwater prawns. In areas of low predation, males tend to have many bright, colorful spots that are attractive to females but are also conspicuous to predators. Spot number, size, and brightness are inherited; the offspring of brightly colored males are brightly colored. In a series of experiments, fish from several populations were exposed to different levels of predation over several generations or transferred between areas with high and low predation intensity. There was a regular decline in spot number, spot size, and spot brightness in populations subjected to more predation both in the field and lab (Endler 1980, 1983). Even the pattern of size-specific predation affected heritable
Where Guppies were exposed to cichlids, which tend to prey chiefly on larger Guppies, Guppies matured later at larger sizes, whereas where the predator was the topminnow, which targets predominantly smaller Guppies, maturation occurred earlier at smaller sizes (Reznick et al. 1990, 1997).
As was pointed out with respect to competition, nonnative predators have unequivocal impacts on native fishes wherever they are introduced, having eliminated populations and even species in many locales. Examples abound (see Introduced predators), but strongest impacts have come from species introduced to improve sport fishing (e.g., Flathead Catfish, Brown and Rainbow trout, Smallmouth and Largemouth black bass, Peacock Cichlid, Nile Perch, Northern Pike and Pikeperch, and even Mosquitofish) (Fuller et al. 1999; Rahel 2002; Helfman 2007).
Related Topics