Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Individuals, populations, and assemblages
Life histories and reproductive ecology of Fishes
Individuals
Ecological adaptations are traits of an individual that insure its survival and reproduction in response to selection pressures from the biotic and abiotic environment. It is important to emphasize the individual as the basic unit of adaptation since natural selection operates primarily at the level of the individual, favoring individuals of one genotype while selecting against individuals with less favorable genotypes. We can then ask if survival and reproduction are enhanced by how an individual selects an appropriate habitat in which to live (discussed below, under Assemblages), how it budgets its time and energy among the activities and conflicting demands presented to it on a daily basis, and how it eventually partitions energy into growth versus reproduction.
Life histories and reproductive ecology
A life history can be viewed as how an individual divides up its time and resources among the often-conflicting demands associated with maintenance, growth, reproduction, mortality, and migration. Life history characteristics or traits are measurable aspects of an individual’s life history and include age- and size-specific birth rates (and associated characteristics such as clutch size, egg size, offspring provisioning, and clutch frequency), and the probabilities of death and migration (Congdon et al. 1982; Dunham et al. 1989). These traits vary among species, among populations within a species, and among individuals and sexes within a population in ways that make evolutionary sense, indicating their adaptiveness. The challenge to biologists is to identify trends in life history traits, identify the likely selection pressures causing variation, and interpret the adaptiveness of the variation (Potts & Wootton 1984; Stearns 1992; Winemiller & Rose 1992; Matthews 1998). Many life history traits are correlated, which means they are inherited together and change in direct relationship with one another, making it somewhat difficult to isolate the exact interaction between environment and adaptation. Nonetheless, their importance in determining reproductive success is obvious.
Analyses of life history traits focus on females, in part because female reproductive effort produces eggs, each of which has a much greater likelihood of becoming a new individual than is the case for the millions of sperm produced by a male. About one dozen life history characteristics (termed “traits” by some but not all authors) have direct links with reproduction and have been identified and quantified in many fishes.
1 Age and size at maturation. A complex but fascinating trade-off exists between early versus late maturation; the trade-off depends on the probability of successful reproduction versus the risk of death. A female that delays maturation until she is larger and older will produce more eggs at each spawning but runs the risk of dying before she ever reproduces (Fig. 24.1). A fish that spawns at an earlier age stands a greater chance of getting some genes into the next generation at least once. However, younger fish are smaller and hence produce fewer and often smaller eggs, which lessens the chance that any will make it past egg and larval predators and starvation. Also, by allocating energy to reproduction, the earlier spawning fish has slower somatic (body) growth and is then more subject to predation because of smaller body size (Werner et al. 1983). Additionally, reproduction uses up much energy, potentially placing a smaller fish with lesser energy stores in a weakened condition, which reduces the chances of future reproduction. Theoretically, females in populations where adult survival is poor should reproduce at an earlier age than in populations where survival is better. Female Guppies in downstream locales in Trinidad where predators are abundant do mature earlier than upstream populations with fewer predators (Reznick & Endler 1982). Similarly, individuals in commercially exploited fish populations, particularly those where adults are targeted by the fishery, often reproduce at earlier ages than do fish in unexploited populations (e.g., O’Brien et al. 1993).
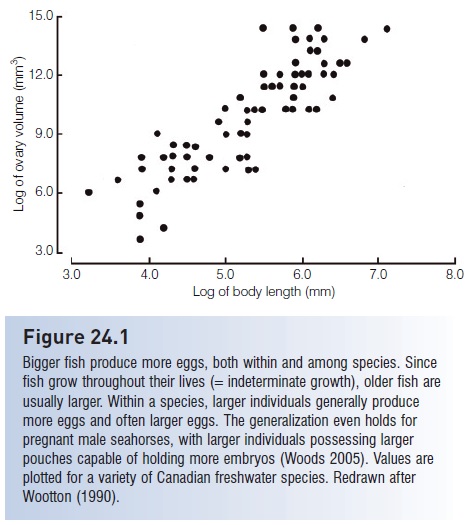
Figure 24.1
Bigger fish produce more eggs, both within and among species. Since fish grow throughout their lives (=indeterminate growth), older fish are usually larger. Within a species, larger individuals generally produce more eggs and often larger eggs. The generalization even holds for pregnant male seahorses, with larger individuals possessing larger pouches capable of holding more embryos (Woods 2005). Values are plotted for a variety of Canadian freshwater species. Redrawn after Wootton (1990).
2 Body size. Even very large predators eat relatively small prey, but only large predators can eat large prey. Therefore, larger fishes are susceptible to predation by fewer predators while at the same time larger fish can catch and swallow a broader range of prey types. Larger fishes are also able to store more energy, to swim faster and farther, and to better overcome harsh abiotic conditions such as strong currents (Karr et al. 1992). Size determines territorial interactions as well as male mating success in many fishes. Also, larger fish emerge from winter with greater energy stores and in better condition than do smaller individuals (Cargnelli & Gross 1997), and larger fish produce eggs with higher hatching success and higher larval survival (Trippel 1995). This premium on large size comes, however, at a cost because energy allocated to somatic growth is unavailable for immediate reproduction, as discussed above.
3 Longevity. The longer an individual lives, the more reproductive opportunities it should have, discounted by how long it waits until first reproduction (point 1 above) and how long an interval exists between reproductive periods (point 10 below).
4 Clutch size. How many eggs a female produces at each spawning varies as a function of body condition and size, age, egg size, and number of spawnings per season (egg number is referred to as fecundity and can be subdivided into batch, breeding season, or lifetime fecundity, although fecundity commonly refers to the number of eggs or young produced per year; Eggs and sperm). Combining clutch size with egg size (point 5 below) gives a measure of reproductive allotment, which is the percentage of a female’s weight devoted to eggs or embryos. Theoretically, females in populations where adult survival is poor should devote more energy to reproduction than in populations where survival is better. Reproductive allotment in female guppies in predator-dense populations is 30% greater than in females subject to less predation (Reznick & Endler 1982). Commercially exploited species (e.g., pike, halibut) show increased fecundities as compared to unexploited populations, a change in a life history trait which could compensate for high levels of predation or exploitation (Policansky 1993b).
5 Egg size and size at birth. Both the mean and range of egg size vary within species and within individuals (Matthews 1998; Wootton 1999). Eggs spawned early in a season for multiple spawners tend to be larger. In the Least Killifish, Heterandria Formosa (Poeciliidae), the more broods a female produces, the smaller the young from each brood will be. In the Orangethroat Darter, Etheostoma spectabile (Percidae), offspring hatched from larger eggs are larger and are less likely to starve (Marsh 1986). Greater investment in each egg, largely in terms of amount of yolk, increases the chances of survival for that offspring. A larger larva is better able to avoid predators and feed independently (e.g., Richards & Lindeman 1987). The volume of the ovarian space in a female determines fecundity, producing an inverse relationship between egg size and number of eggs. Fish that produce larger eggs have lower batch fecundities.
6 Time until hatching and exogenous feeding. For egglaying species, which is most fishes, eggs are deposited on substrates or in the water column and are essentially defenseless, either immobile or floating. After hatching, yolk-sac fry are inefficient swimmers. The longer a larva spends growing inside the egg or absorbing yolk resources, the larger it will become before having to obtain its own food. A larva trades off the increased vulnerability it experiences while being passive against the advantages it will have in finding food and avoiding predators once it achieves independence from the egg shell and yolk sac.
7 Larval growth rate and interval length. Rapid growth provides a larva with the same advantages as large egg size or yolk supply, namely achieving a larger size earlier. However, rapid growth requires more energy and higher metabolism, which in turn demands more efficient or faster feeding and an increased likelihood of starvation. A short larval period means larvae can transform quickly into juveniles, and settle from the plankton and into the generally safer juvenile habitat. But if a larva finds itself in an inappropriate habitat at the end of its larval period – such as far out at sea for a species adapted to shallow water existence as a juvenile and adult – then a short larval life provides little advantage (but see below, Synthesis: what determines assemblage structure among coral reef fishes?). Conversely, a long larval interval can permit long-distance dispersal. However, extended planktonic life exposes the larva for a longer time to the extreme hazards of planktonic existence, when >99% of larvae are eaten or starve. Growth rates of juveniles and adults are subject to advantages and constraints as discussed in point 2.
8 Spawning bouts per year and duration of spawning season. The number of times an individual, particularly a female, spawns each year tells much about the allocation of energy to reproduction. Duration of the spawning season is more a population than an individual characteristic and is useful in assessing potential recruitment into that population.
9 Number of spawnings per lifetime. Most fishes are iteroparous (itero =to repeat, as in reiterate; parous =to give birth), spawning repeatedly throughout their lives. One-time spawners, termed semelparous, devote all their energy to a single, massive spawning event, after which they die (anguillid eels, many Pacific salmons, lampreys, some gobies).
10 Reproductive interval. The time spent between reproductive bouts for iteroparous species varies greatly, from daily spawners that reproduce yearround in some low-latitude coral reef fishes (e.g., wrasses), to fishes that spawn every few weeks during a protracted season (e.g., Grunion, darters), to seasonal spawners that may spawn only once or a few times during a limited season (snappers, groupers, larger percids, centrarchid basses), to internal bearers with long gestation periods of a year or more (some sharks), to species that may wait several years between spawnings (sturgeon). For iteroparous species that spawn repeatedly each year, the reproductive interval can theoretically be adjusted in response to expected mortality levels. Where the probability of mortality is high, reproductive intervals should be short. As was the case for variable age at first reproduction and reproductive allotment (see above), female guppies exposed to high levels of predation
have relatively short reproductive intervals (Reznick & Endler 1982).
11 Parental care. The degree of care given has an overwhelming influence on the mortality rate of the young and is generally inversely proportional to fecundity (see Parental care). Parental care is often distinguished as prezygotic (e.g., nest preparation) and postzygotic (e.g., internal brooding, guarding young). Care may be nonexistent to rather elaborate. Broadcast spawners release large numbers of eggs into the water column and provide no further care (tarpon, cods, tunas). Moderate care occurs in fishes that spawn intermediate numbers of eggs on substrates and may involve some substrate preparation such as nest construction or egg covering (salmons, Grunion). More extensive care occurs in fishes that prepare a nest and then guard relatively few eggs until they hatch and perhaps a little later (sticklebacks, sunfishes, some cichlids). Intensive care is usually associated with relatively low numbers of large eggs, such as fishes that gestate young internally (livebearers, embiotocid surfperches) or incubate the eggs orally (cardinalfishes); some oral incubators continue to protect the young after hatching (some catfishes) and some cichlids even feed their young with external body secretions. Male Bowfin (Amia calva) are egg layers without mouth brooding that guard their young until they are several centimeters
long, a rarity among pre-teleosteans and teleosts alike. Parental care increases survival of the young but occurs at a cost to the parents because extended care increases the interval between spawnings.
12 Gender change and sex ratio variation. Fishes in several families change sex, beginning as males and changing to females (protandry) or vice versa (protogyny) . The timing of the change is largely determined by the relative reproductive success males or females experience at the same body size. Sex change occurs at a cost in immediate reproductive output, because converting the actual machinery of gamete production from one sex to another may require weeks or months. In many vertebrates (crocodilians, turtles, lizards, possums, monkeys), the sex of offspring may be determined by conditions such as the temperature at which the eggs or embryos develop (=environmental sex determination, ESD). Extreme temperatures affect sex determination in a few fishes, mostly atheriniforms such as rivulins, ricefish, and livebearers; pH can also influence sex determination in some cichlids and a livebearer (Rubin 1985; Francis 1992). Naturally occurring variation in temperature determines the sex of developing Atlantic Silversides, Menidia menidia
(Atherinidae). Offspring produced early in the year at relatively low temperatures tend to be female, whereas young produced later at higher temperatures tend to be male (see Determination, differentiation, and maturation). Females could
theoretically manipulate the sex ratio of their offspring to take advantage of disproportionate numbers of one sex or of environmental conditions that favor one sex over another. Whether and why a female silverside actively manipulates the sex ratio of her young is a matter of conjecture. It is difficult to imagine how a female would assess current sex ratios; in addition, larvae are dispersed widely and are unlikely to be recruited into the same population and environment as the mother (Conover & Kynard 1981; Conover & Van Voorhees 1990; D. Conover, pers. comm.).
13 Geographic patterns and phylogenetic constraints. In many families, related species living in different habitats often adopt life history patterns appropriate for that habitat, and unrelated fishes converge on suites of life history adaptations. Mouth-brooding fishes worldwide have converged on small clutches of large eggs, slow growth rates, and protracted breeding seasons (bonytongues, marine catfishes, cichlids). Such convergence is evidence of the importance of environmental selection factors promoting one life history over another and can be found at relatively large geographic scales. Among freshwater fishes in North America, species that mature relatively late in life tend to have larger body sizes, longer life spans, higher fecundities, smaller eggs, few multiple spawnings, and a short spawning season (sturgeons, Paddlefish, shads, muskellunge, charrs, Burbot). Fishes with extended spawning seasons tend to have larger eggs, multiple spawning bouts, and exhibit more parental care (cavefishes, madtom catfishes). Marine fishes that have extensive geographic ranges (tarpon, cods) also tend to have high fecundity. Anadromous species, such as salmons, Striped Bass, and sturgeons, mature late, grow fast as adults, live long, and have large eggs (Winemiller & Rose 1992). As with any evolved characteristic, life history traits are influenced by the evolutionary history of the lineage to which an animal belongs. Consequently, an animal may not have the life history characteristics that we expect given current conditions. Unless selection pressures have been relatively stable for many generations, an animal’s adaptations will not necessarily reflect present conditions but will instead reflect past adaptive scenarios and selection pressures. For relatively conservative traits that are shared among many members of a lineage, historical constraints may be difficult to overcome and species will retain seemingly nonadaptive or nonoptimal characteristics. Regardless of latitude and habitat, percopsiforms (Troutperch, Pirate Perch, cave fishes) tend to be small, produce small clutches of large eggs, exhibit extensive parental care, and have protracted spawning seasons and slow growth rates. Within the cypriniforms, suckers in the genus Ictiobus are large with large clutches and few spawning bouts whereas minnows in the genus Notropis are small, have small clutches, and frequent spawning bouts. Flatfishes as a group mature at large size, produce large clutches of small eggs during short spawning seasons, and grow rapidly when young (Winemiller & Rose 1992).
Related Topics