Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Individuals, populations, and assemblages
Habitat use and choice - Diversity of Fishes: Assemblages
Habitat use and choice
An important component of a species’ niche, and one that can easily differ among species, is its habitat. Habitat can often be described and quantified in detail. Among stream fishes, species often differ in height above bottom (Fig. 24.5), preferred current strength, bottom type (particle size and type), structure, distance from shore, amount of vegetation, and type and amount of food resources. A survey of major habitats in an eastern North American stream (rapids, riffl es, runs, pools, overhangs) often shows that species segregate along vertical dimensions, with certain species typically found in contact with the bottom (catfishes, darters, sculpins, eels, grazing minnows), some just above the bottom (suckers), some species low and others higher in the water column (planktivorous minnows, trout), some close to the surface (silversides, livebearers, topminnows), some in swift water (darters, trout), and others in moderate flow or in slow flow near more rapidly flowing water (catfishes, minnows, pickerel, sunfishes). The existence of particular species in particular habitats implies an active choice by individuals (e.g., Gorman 1988). Experimental studies, usually of juveniles, generally show that individuals actively choose habitats, that the ones they prefer are the ones in which the species is most often found, and that preferred habitats are ones in which a species can successfully feed, avoid predators, and reproduce, i.e., habitat choice is an evolved aspect of a species’ niche (e.g., Sale 1969). Habitat choice is, however, dynamic within a species, varying on the basis of age, size, sex, reproductive condition, geographic area, and environmental conditions (e.g., Karr et al. 1982; Allan & Castillo 2007).
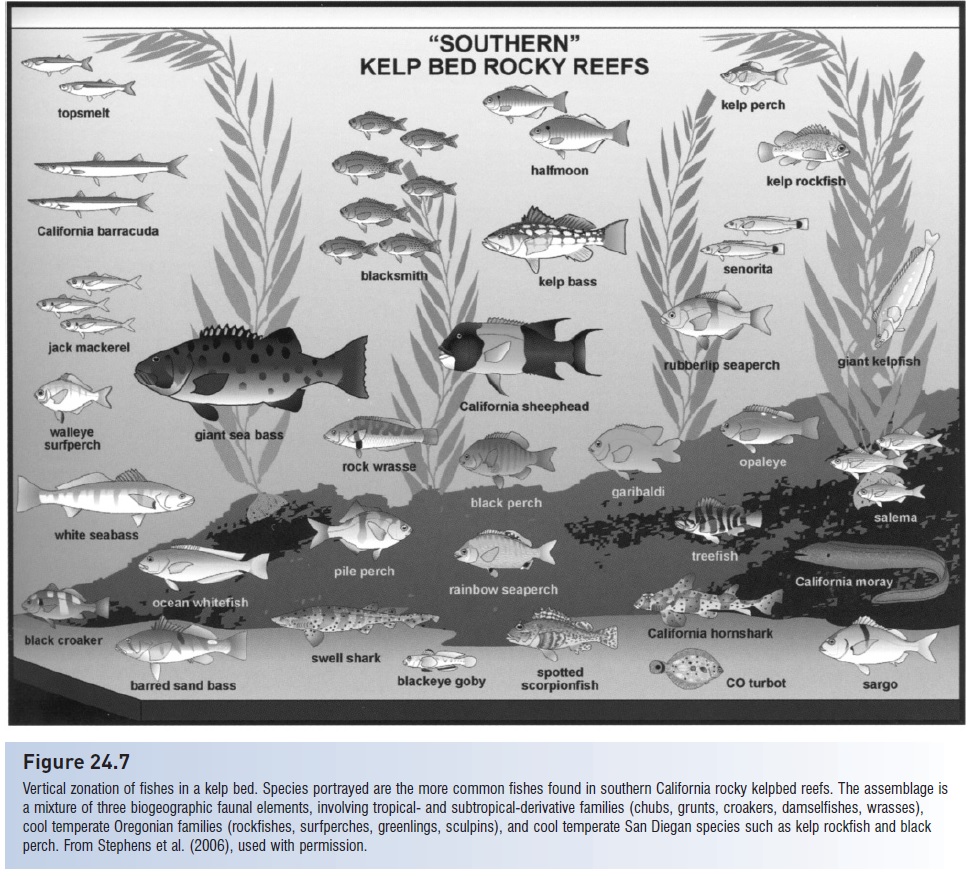
Similar descriptions, based on habitat characteristics, apply to assemblages in most major habitat types. Different faunas in different geographic locales often occupy similar habitats and, in essence, converge on many niche characteristics. The Chinese Algae Eater,Gyrinocheilus (Gyrinocheilidae), belongs to a Southeast Asian family related to loaches. It lives in rivers where it feeds on algae attached to rocks. It has a sucking mouth with which it clings to
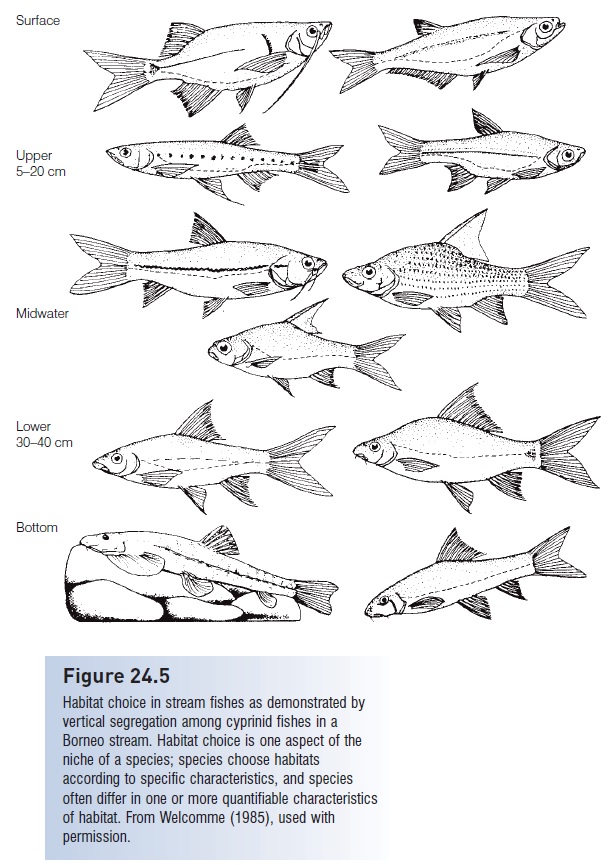
Figure 24.5
Habitat choice in stream fishes as demonstrated by vertical segregation among cyprinid fishes in a Borneo stream. Habitat choice is one aspect of the niche of a species; species choose habitats according to specific characteristics, and species often differ in one or more quantifiable characteristics of habitat. From Welcomme (1985), used with permission.
A South American catfish, Otocinclus (Loricariidae), similarly feeds on algae-covered rocks in flowing water. It is convergent with the Algae Eater in body form and size, suctorial mouth, and even coloration (see Strong currents and turbulent water). Stream-dwelling galaxiids of the southern hemisphere, especially in New Zealand and Australia, occupy niches very similar to those of salmonid trouts in the northern hemisphere. They are convergent in body morphology, habitat, and foraging habits and have been rapidly exterminated by the introduction of trout in many locales due to competition and predation. A characin, the Dorado, Salminus maxillosus, is a desirable gamefish that lives in streams of the Amazon region (Fig. 24.6). Its morphology, coloration, and piscivorous foraging habits are, as its generic name implies, remarkably convergent with stream-dwelling salmonids such as Brown Trout (Esteves & Lobo 2001).
A very broad and striking convergence occurs among approximately 60 families of fishes that fill the eel niche in their respective assemblages. Only 22 of these families belong to the order Anguilliformes and are “true” eels. Many of these fishes are convergent in habitat choice, occurring on and often in the bottom in soft sediments or among the interstices of rocks and other structure. They share other characteristics: elongated dorsal and anal fins that often lack hard spines, increased vertebral counts,

Figure 24.6
The characid Dorado, a popular gamefish of the Amazon Basin, is ecologically convergent with large riverine salmonids. Photo from Rio Paraná, Ayollas, Paraguay, courtesy of www.planetapesca.com.
Behaviorally they are carnivores and scavengers (some are parasitic), can swim backwards and forwards with equal facility (palindro mic locomotion), and tear pieces off their prey by holding on and rotating rapidly along their long body axis.
Habitat choice changes with the seasons, with size and age of fish, and also with the presence of other species, particularly predators and competitors. Many stream species, such as darters and benthic minnows and suckers, occupy increasingly swift water as they grow older. Often the distribution results from spawning habits: adults migrate upstream to spawn in headwaters and young move progressively downstream with growth (e.g., Hall 1972). Such an ontogenetic habitat shift could reflect a body size constraint related to the current speed at which an individual can hold position without expending excess energy or using too much oxygen (see below and Fig. 5.7). A general and somewhat unexplained pattern of increasing depth with age occurs in a number of freshwater and marine species. In many species, this trend reflects the use of inshore, shallow, productive nursery areas that are reputed to be relatively predator-free (e.g., saltmarshes, mangroves); these fishes move offshore as they grow. In other species (minnows, copper sweepers, damselfishes, surgeonfishes, sunfishes, croakers, percids, clinids, wrasses, Great Barracuda), an actual size–depth correlation has been found (Helfman 1978; Power 1987). That the relationship exists in a diversity of habitats and involves a number of unrelated species (e.g., Polloni et al. 1979) implies a convergent, adaptive trait, but the details of this adaptation remain to be worked out in most cases.
“Bigger–deeper” distributions within species and other types of habitat shifts may result from avoidance of predators, influenced by differences in vulnerability between different size classes of prey.
When predatory sunfishes (Lepomis spp.) are present and Largemouth Bass are absent, young-of-the-year Stoneroller Minnows (Campostoma anomalum) occupy shallows of pools, whereas larger Stonerollers prefer deeper portions. Sunfishes occupy deeper portions of pools and can eat young minnows but are too small to eat larger minnows. When bass, which can eat all minnows and also prefer deep sections, are added, all size classes of Stoneroller are confined to shallow margins or emigrate from pools.
A combination of fish, avian, and mammalian predators affect the depth distributions of loricariid catfishes (including the “plecostomus” species of the aquarium trade) in Panamanian streams. These herbivores feed on algae that grow most abundantly in the shallows of pools. Small catfishes live in the more productive shallows, whereas large catfishes occur in deeper water where algal production is minimal and where the fish lose fat reserves and cease growing. The habitat differences of the two size groups are enforced by the balanced impact of terrestrial predators and piscivorous fishes. Birds and mammals can capture any size prey whereas piscivorous characins are gape-limited and cannot swallow the large catfishes. Large catfishes are therefore forced into deeper, less productive regions by terrestrial predation. The small catfishes avoid predatory fishes that live in deep water but can hide from the birds and mammals in shallow water by seeking refuge among rocks; however these refuges are too small for larger catfishes. Cannibalism can also influence habitat shifts, forcing smaller fish into suboptimal habitats where their growth rates suffer. Smaller sculpins (Cottus) use a diversity of habitats in some streams and prefer deeper water in others when large conspecifics are missing. If larger fish are present or are added experimentally, the smaller fish shift to shallower habitats. Similar habitat shifts, resulting in the occupation of suboptimal feeding habitats by small prey, have been demonstrated in other minnow species and Bluegill Sunfish (Werner et al. 1983; Gilliam & Fraser 1987; Schlosser 1987; Freeman & Stouder 1989).
Related Topics