Chapter: Genetics and Molecular Biology: Chemotaxis
Phosphorylation and the Rapid Response
Phosphorylation and the Rapid Response
Chemotaxis requires sensing an extracellular
condition and then signaling to some intracellular component. A number of
other systems, for example the phosphotransfer system, the nitrogen regulatory
system, phosphate uptake system, and porin system, use similar signaling
path-ways. These pathways contain sensor proteins which often span the membrane
and serve to couple extracellular conditions to internal activity. The sensors
are similar in structure to one another, phospho - rylate themselves on
histidine residues, and phosphorylate the regulator proteins on aspartate
residues. The regulator proteins also are similar in structure to one another.
In order that the stimulation be reversible,
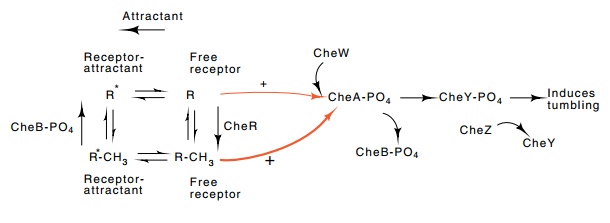
Figure
22.13 Pathway of methylation and
phosphorylation. The methylatedstate of free receptor is most active in
stimulating phosphorylation of CheA. CheB phosphate demethylates the receptor
and CheR methylates it.
In the chemotaxis system the Tar, Tsr, Trg, or Tap
proteins transmit a signal across the inner membrane. These signals are
received by the CheW and CheA proteins and are translated into varying levels
of phosphorylation of CheA. This was found by analogy to the phos-photransfer
system and a finding by Magasanik that phosphorylation was involved with
regulation of nitrogen metabolism. When the purified chemotaxis proteins are
appropriately incubated together in vitro,
a phosphate is transferred from ATP to CheA protein. To demonstrate that this
pathway is involved with chemotaxis, mutants lacking the histidine to which the
phosphate is transferred have been found not to be chemotactic.
Two further phosphorylation pathways diverge from
CheA. The first is phosphorylation of CheY from CheA. This is a rapid step that
leads to signaling the flagellar motor, most likely by direct interaction
be-tween phosphorylated CheY protein and the motor. This reaction in-duces
tumbling. Phosphorylated CheA also leads to phosphorylation of CheB, which
demethylates the receptors.
The response of the chemotaxis pathway involves the complicated interaction of many components (Fig. 22.13). An increase in the con-centration of attractant rapidly decreases the phosphorylating signal from receptor to CheA-CheW, and as a result decreases the concentra - tion of phosphorylated CheY. Tumbling is suppressed and smooth swimming is prolonged. Additionally, the decrease in phosphorylation of CheA decreases the phosphorylation of CheB which decreases its demethylation activity. As a result, but one taking place more slowly than the first response, methylation of the receptor increases, the cells begin to adapt, and the phosphorylation levels begin to increase. Overall, then, the phosphorylation levels respond rapidly and transmit a signal to the motor, and the methylation levels respond more slowly and serve to bring the phosphorylation levels back to the levels characteristic of the absence of attractant.
Chemotaxis is remarkable. A single cell manages to
sense attractants with receptors located outside the cytoplasm, transmit the
information to the interior, remember the information it has received so as to
adapt if the attractant or repellent concentration is no longer changing, build
and energize a motor, and synthesize a bundle of flagella that smoothly rotate
together or fly apart when necessary. The biochemical bases for parts of the
system are known. A few of the receptors have been purified, and the center of
the adaptation phenomenon, the methylation and phosphorylation of the proteins,
is partially understood. Whether we will be able to understand the underlying
physical chemistry of the protein conformational changes and the drive
mechanism of the flagel-lar motor in the years ahead is an interesting
question.
Related Topics