Chapter: Genetics and Molecular Biology: Chemotaxis
How Cells Swim
How Cells Swim
Escherichia
coli propels itself through liquid by
rotating its flagella. Normally the flagella are left-handed helices and their
rotation generates a thrust that moves the cell. In this section we consider
the structure of flagella, how it’s known they rotate, how the rotation is
created, and how the several flagella present on a single cell can function
together.
Flagella are too thin to be easily seen by ordinary
light microscopes, but they can be visualized with light microscopes operating
in the dark-field mode, interference microscopes, video processing, or
elec-tron microscopes. Careful isolation of flagella shows that they are
attached to a hook-shaped structure connected to a set of rings that is
embedded in the cell’s membranes. The rings have the appearance of a motor that
rotates the flagella (Fig. 22.4). The hook is a flexible connec-tor between the
basal structure and the flagella. Such a universal joint
Figure
22.4 The appearance of a flagellum at
low magnification in the electronmicroscope and the structure of the basal
body, the motor, at high magnifica-tion showing the rings and the membranes of
the cell wall.
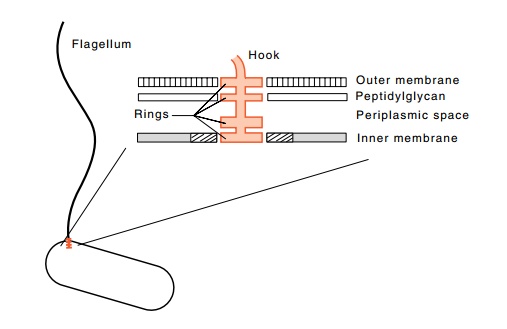
is necessary because in E. coli the flagella
sprout from random points on the cell’s surface and point in several different
directions. A cell typically contains about six flagella. These must join
together in a bundle in a way that permits each to rotate in response to its
motor. The hook acts as a universal joint that permits the torque to be
transmitted around a bend.
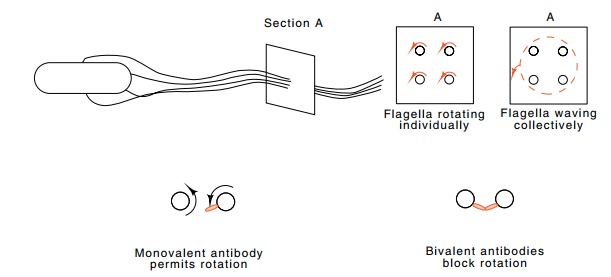
Figure 22.5 A bacterium with flagella sprouting from various locations bend-ing via the hook portion and coming together in a bundle. The section taken at point A shows flagella rotating individually and their movement if the bundle waves as a whole.
Because of the size of the flagella, indirect means
must be used to demonstrate that they rotate. One simple experiment uses the
observa-tion that antibody against flagellin can block motility. More
precisely, bivalent antibody blocks motility, but monovalent antibody does not
(Fig. 22.5). This result can be understood if the flagella form a bundle
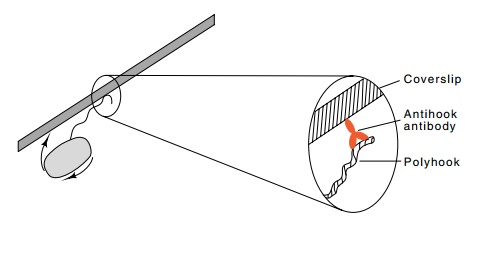
Figure
22.6 Demonstration of the hook’s
rotation. After attachment of thehook or a short flagellum to a cover slip, the
cell rotates, and this can easily be observed.
and each flagellum rotates within this bundle. A
bivalent antibody molecule can link different flagella and prevent their
rotation, but if flagella waved or rotated as a group, bivalent antibodies
would have an effect no different from monovalent antibodies.
The most graphic demonstration that flagella rotate
is also the basis of many other important experiments on chemotaxis. Simon used
one mutation to block flagellin synthesis and another to permit greater than
usual growth of the hook. As the synthesis of flagella is sensitive to
catabolite repression, growth of cells in glucose reduced the number of the
resulting polyhooks from about six per cell to about one. These cells could be
bound to a microscope slide by means of antihook antibody that had bound to the
hook and nonspecifically bound to the glass as well (Fig. 22.6). Chemotactic
cells immobilized in this way rotated at two to nine revolutions per second.
This leads to the conclusion that the hook normally rotates, but when it was
fastened to the microscope slide instead, the cell rotated. Nonmotile cells did
not rotate. Of course, motile but nonchemotactic mutants did rotate because
they can swim but do so in a nondirected fashion and are incapable of swimming
up a gradient.
The immobilization experiment shows that a single
flagellum rotates and that the rotation is associated with chemotaxis.
Dark-field micros-copy has shown that the bundle of flagella on a cell is
stable as long as the flagella rotate counterclockwise. If the flagella reverse
their rotation, their left-helical structure compels the bundle to fly apart
temporarily. Furthermore, if the reversal is sufficiently vigorous, the
flagella snap into a right-helical conformation. This further ensures that the
bundle of flagella disperses.
Related Topics