Chapter: Genetics and Molecular Biology: Chemotaxis
Energy for Chemotaxis
The Energy for Chemotaxis
Application of genetic and biochemical methods has
allowed determination of the energy source required for chemotaxis. On one
hand it seems logical that ATP would be the direct source of the mechanical
energy required for swimming since most energy transductions in higher cells
appear to use ATP. On the other hand, the flagella originate in the cell
membrane, and a substantial proton gradient exists across the inner membrane
under most growth conditions. Therefore the direct source of energy could also
derive from the membrane potential.
ATP and the proton motive force across the inner
membrane are normally interconvertible by means of the membrane-bound ATPase
(Fig. 22.9). The membrane potential generates ATP and, conversely, ATP can be
used to generate a membrane potential. Therefore, blocking formation of either
ATP or the membrane potential could have an effect on the other. Arsenate
permits decoupling to determine what actually
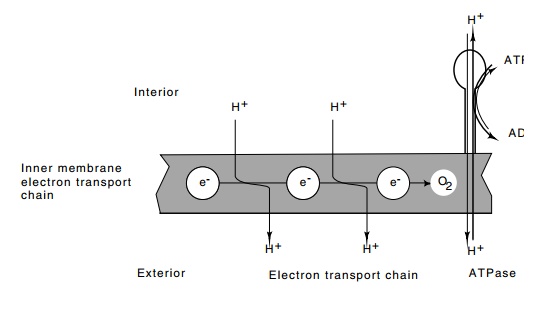
Figure
22.9 A schematic of the inner membrane
of a cell showing that electronflow down the electron transport chain leads to
export of protons and ultimate transfer of electrons to oxygen. Reentry of protons
to the cell through the ATPase generates ATP. Conversely, ATP can be hydrolyzed
by ATPase to pump protons out of the cell.
drives the motor of the flagella. It blocks ATP
formation both by glycolysis and by ATPase, but does not directly affect the membrane
potential. Cells treated with arsenate are found to be motile, but they do not
swim up gradients of attractants. This shows that ATP is not required for
motility, but probably is involved with the process of modulating run duration.
Next, to investigate the role of the membrane
potential, cells can be grown anaerobically. This blocks the usual means of
generation of the membrane potential because the electron transfer chain
becomes inac-tive owing to the lack of a terminal electron acceptor. Then, to
prevent energy from ATP from being used to create a membrane potential, an
ATPase-negative mutant can be used. Anaerobically grown ATPase mutants are
found not to be motile, leading to the conclusion that the motors that drive
the flagella are run by the cell’s membrane potential. Other experiments show
that cells swim when transmembrane pH gradient or membrane potentials are
generated artificially.
Related Topics