Chapter: Genetics and Molecular Biology: Oncogenesis, Molecular Aspects
Bacterially Induced Tumors in Plants
Bacterially Induced Tumors in Plants
Plants appear to be simpler than mammals and yet
they are susceptible to tumors just like mammals. Therefore, some of the
lessons that we learn about plant tumors may be helpful in the analysis of
animal tumors. One feature that makes the study of plant cells and plant tumors
valuable in research on oncogenesis is that cell cultures from some plants can
be maintained indefinitely in cell culture, but when desired, these
undifferentiated cells can be induced to differentiate back into normal plants
that reproduce sexually. Such techniques permit detailed analysis of the causes
of oncogenesis. In addition, these approaches have the potential for yielding
valuable mutants by using techniques similar to those used with bacteria for
the isolation of single mutant cells from large cultures and then regenerating
complete mutant plants.
The bacterium Agrobacterium
tumefaciens can induce the growth of masses of undifferentiated cells,
called crown galls, in susceptible plants. This transformation to the
undifferentiated state requires a 200-Kb plasmid carried by the bacterium. In
the transformation process at least 8 to 10 Kb of DNA from the plasmid are
transferred from the bacterium into the plant cells. There the DNA is
integrated into the chromosome of the plant, where it is replicated along with
the cellular DNA. As a result, all cells of a crown gall contain fragments of DNA
originating from the plasmid. Part of the integrated plasmid DNA directs the
plant to synthesize and secrete the compounds octopine or nopaline (Fig. 23.1).
In turn, these compounds can be catabolized by the Agrobacter bacteria in the crown gall. Few other bacteria or
parasites can utilize these compounds for growth. Thus, one Agrobacterium
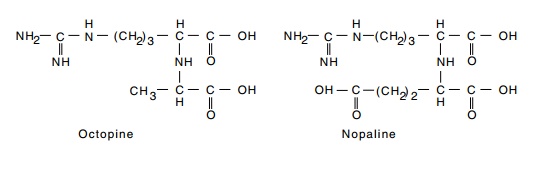
Figure 23.1 Structures of octopine and nopaline.
In contrast to the nontransformed plant cells, the
cells from crown galls do not require the growth factors auxin and cytokinin
for their continued growth in culture (Fig. 23.2). Ordinarily, medium for plant
cells requires the presence of these two small-molecule growth factors in
addition to a variety of other metabolites. With the ratio of auxin and
cytokinin at one value, plant cells in culture remain undifferentiated, but if
the ratio of auxin to cytokinin is increased, stems and leaves tend to develop.
Conversely, if the auxin to cytokinin ratio is decreased, the cells become
root-like. Without either auxin or cytokinin present, the cells do not grow. In
the whole plant, auxin is synthesized in the stem tips and cytokinin in the
root tips. A concentration gradient in these molecules from the top to bottom
of the plant helps cells identify their positions and develop appropriately.
Plant cells transformed with Agro-bacter do
not require either auxin or cytokinin for growth. It is likelythat the DNA that
was acquired from the plasmid by the transformed cells directs or induces
synthesis of auxin and cytokinin-like substances that substitute for these
chemicals both in the crown gall and in cell culture. By analogy to the crown
galls in plants, it was predicted and found that many types of animal cancers
would involve alterations in the cell’s synthesis of, or their response to,
growth factors.
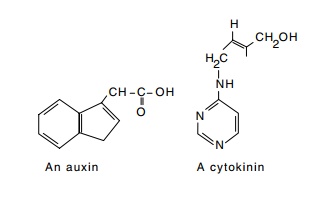
Figure 23.2 Indole 3-aceticacid, an auxin and 6-(4-hy-droxy-3-methyl-trans-2-buten-ylamino)purine, a cytokinin.
The DNA transfer mechanism utilized by Agrobacter can be utilized for genetic
engineering. The DNA to be introduced to the plant cell can be included between
the DNA recombination sequences of the transfor-mation plasmid and
transformation then occurs much like lambda phage integration.
Related Topics