Chapter: Basic & Clinical Pharmacology : Adrenoceptor Agonists & Sympathomimetic Drugs
Noncardiac Effects of Sympathomimetics - Organ System Effects of Sympathomimetic Drugs
Noncardiac Effects of
Sympathomimetics
Adrenoceptors
are distributed in virtually all organ systems. This section focuses on the
activation of adrenoceptors that are responsible for the therapeutic effects of
sympathomimetics or that explain their adverse effects. A more detailed
description of the therapeutic use of sympathomimetics is given later.
Activation
of β2 receptors in bronchial
smooth muscle leads to bronchodilation, and β2 agonists are important in the treat-ment of
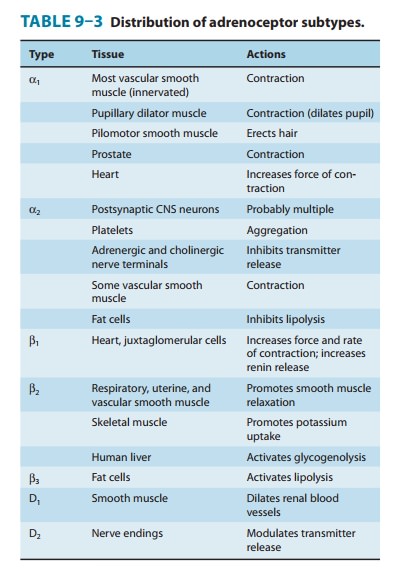
asthma (see Table 9–3).
In the eye, the radial pupillary dilator muscle of the iris con-tains α receptors; activation by drugs such as phenylephrine causes mydriasis (see Figure 6–9). Alpha stimulants also have important effects on intraocular pressure. Alpha agonists increase the outflow of aqueous humor from the eye and can be used clinically to reduce intraocular pressure. In contrast, β agonists have little effect, but βantagonists decrease the production of aqueous humor. These effects are important in the treatment of glaucoma , a leading cause of blindness.
In
genitourinary organs, the bladder
base, urethral sphincter, and prostate contain α receptors that mediate contraction and
therefore promote urinary continence. The specific subtype of α1 receptor involved in
mediating constriction of the bladder base and prostate is uncertain, but α1A receptors probably
play an important role. This effect explains why urinary retention is a
potential adverse effect of administration of the α1 agonist midodrine.
Alpha-receptor
activation in the ductus deferens, seminal vesi-cles, and prostate plays a role
in normal ejaculation. The detumes-cence of erectile tissue that normally
follows ejaculation is also brought about by norepinephrine (and possibly
neuropeptide Y) released from sympathetic nerves. Alpha activation appears to
have a similar detumescent effect on erectile tissue in female animals.
The
salivary glands contain
adrenoceptors that regulate the secretion of amylase and water. However,
certain sympathomi-metic drugs, eg, clonidine, produce symptoms of dry mouth.
The mechanism of this effect is uncertain; it is likely that central ner-vous
system effects are responsible, although peripheral effects may contribute.
The
apocrine sweat glands, located on
the palms of the hands and a few other areas, respond to adrenoceptor
stimulants with increased sweat production. These are the apocrine
nonthermo-regulatory glands usually associated with psychological stress. (The
diffusely distributed thermoregulatory eccrine sweat glands are regulated by sympathetic cholinergic postganglionic
nerves that activate muscarinic cholinoceptors;)
Sympathomimetic
drugs have important effects on intermedi-ary metabolism. Activation of β adrenoceptors in fat cells leads to increased
lipolysis with enhanced release of free fatty acids and glycerol into the
blood. Beta3 adrenoceptors play a role in mediat-ing this response
in animals, but their role in humans is probably minor. Human fat cells also
contain α2 receptors that inhibit lipolysis by decreasing intracellular
cAMP. Sympathomimetic drugs enhance glycogenolysis in the liver, which leads to
increased glucose release into the circulation. In the human liver, the effects
of catecholamines are probably mediated mainly by β receptors, though α1 receptors may also play a role.
Catecholamines in high concentration may also cause metabolic acidosis.
Activation of β2 adrenoceptors by endogenous epinephrine or by sympathomi-metic
drugs promotes the uptake of potassium into cells, leading to a fall in
extracellular potassium. This may result in a fall in the plasma potassium
concentration during stress or protect against a rise in plasma potassium
during exercise. Blockade of these recep-tors may accentuate the rise in plasma
potassium that occurs dur-ing exercise. On the other hand, epinephrine has been
used to treat hyperkalemia in certain conditions, but other alternatives are
more commonly used. Beta receptors and α2 receptors that are expressed in pancreatic
islets tend to increase and decrease insulin secretion, respectively, although
the major regulator of insulin release is the plasma concentration of glucose.
Catecholamines are important endogenous regulators of hor-mone secretion from a number of glands. As mentioned above, insulin secretion is stimulated by β receptors and inhibited by α2 receptors.
Similarly,
renin secretion is stimulated by β1 and inhib-ited by α2 receptors; indeed, β-receptor antagonist drugs may lower blood
pressure in patients with hypertension at least in part by low-ering plasma
renin. Adrenoceptors also modulate the secretion of parathyroid hormone,
calcitonin, thyroxine, and gastrin; however, the physiologic significance of
these control mechanisms is probably limited. In high concentrations,
epinephrine and related agents cause leukocytosis, in part by promoting
demargination of seques-tered white blood cells back into the general
circulation.
The
action of sympathomimetics on the central
nervous sys-tem varies dramatically, depending on their ability to cross
theblood-brain barrier. The catecholamines are almost completely excluded by
this barrier, and subjective CNS effects are noted only at the highest rates of
infusion. These effects have been described as ranging from “nervousness” to
“an adrenaline rush” or “a feeling of impending disaster.” Furthermore,
peripheral effects of β-adrenoceptor agonists such as tachycardia and
tremor are similar to the somatic manifestations of anxiety. In contrast,
noncatecholamines with indirect actions, such as amphetamines, which readily
enter the central nervous system from the circulation,produce qualitatively
very different CNS effects. These actions vary from mild alerting, with
improved attention to boring tasks; through elevation of mood, insomnia,
euphoria, and anorexia; to full-blown psychotic behavior. These effects are not
readily assigned to either α- or β-mediated actions and may represent
enhancement of dopamine-mediated processes or other effects of these drugs in
the CNS.
Related Topics