Chapter: The Massage Connection ANATOMY AND PHYSIOLOGY : Muscular System
Muscle Energetics
Muscle Energetics
The muscle requires energy for contraction to take place and energy is derived from ATP.
ATP → ADP + P +energy
As previously explained, ATP is required for actin- myosin interaction. It is also required for actively pumping calcium into the sarcoplasmic reticulum from the sarcoplasm after the contraction process and maintaining the ionic concentration in the mus-cle fiber by the action of the Na-K ATPase pump lo-cated in the cell membrane. These are only some ac-tivities that require ATP.
It is not possible for the body to have the tremen-dous supply of ATP demanded when a muscle con-tracts. Instead, the body has enough ATP and other high-energy compounds to begin contraction. Typi-cally, a single muscle fiber has enough ATP to support only about 10 twitches or isometric contraction that can last for just 2 seconds; however, it can generate ATP at almost the same rate as the demand through various metabolic processes.
The ATP produced in the muscle fiber at rest is used to transfer energy to another high-energy com-pound, creatine phosphate, present in the cell.
ATP → ADP + phosphate+energy
Creatine+ phosphate +energy → creatine+ phosphate+ ADP
When needed, this compound is broken down by creatine phosphokinase to liberate energy that can be used to form ATP.
Creatine phosphate → creatine+ phosphate+ energy
ADP+ phosphate+ energy → ATP
At rest, a muscle fiber has about six times as much creatine phosphate as ATP. This store is sufficient to produce about 70 twitches or tetanic, isometric con-tractions that last about 15 seconds. If the fiber must sustain its contractions for longer than this, it must rely on other mechanisms for energy. This may be ac-complished by breaking down glucose to lactic acid. Glucose may be made available by the breakdown of stored glycogen or glucose from the blood. All of the above can be achieved without the supply of oxygen.
AEROBIC METABOLISM
The sarcoplasm of the muscle has numerous mito-chondria. The mitochondria have the enzymes neces-sary for breaking down glucose, amino acids, and fatty acids to produce large amounts of ATP in the presence of oxygen. The necessary substrates (e.g., glucose) and oxygen are brought to the muscle by blood. The sarcoplasm also has specialized proteins (myoglobin) that, similar to hemoglobin, combines with oxygen reversibly. In addition, the muscle has some glucose stored in the sarcoplasm in the form of glycogen.
At rest, the energy required by the muscle is pro-vided by aerobic metabolism. The mitochondria ab-sorb the substrate, mostly in the form of fatty acid, ADP, phosphate ions and oxygen from the sar- coplasm and, in the presence of oxygen, forms 17 ATP from each molecule. This process of producing ATP is complex and involves numerous intermediary steps and many enzymes present inside the mito-chondria. This biochemical process is known as the
Krebs cycle or the tricarboxylic acid cycle or TCA cycle, in which the carbon atoms of the substratemolecule are converted to carbon dioxide and the hy-drogen ions generated in the cycle are converted to water.
When the muscle begins to contract, pyruvic acid(derived from glucose) is used as the substrate in the TCA cycle, rather than fatty acid.
ANAEROBIC METABOLISM
Even in the absence of oxygen, the muscle is able to manufacture some ATP (see Figure 4.14). However, few ATP can be produced in this way. Also, the metabolites formed change the pH of the environment and prolonged muscle activity cannot be maintained.
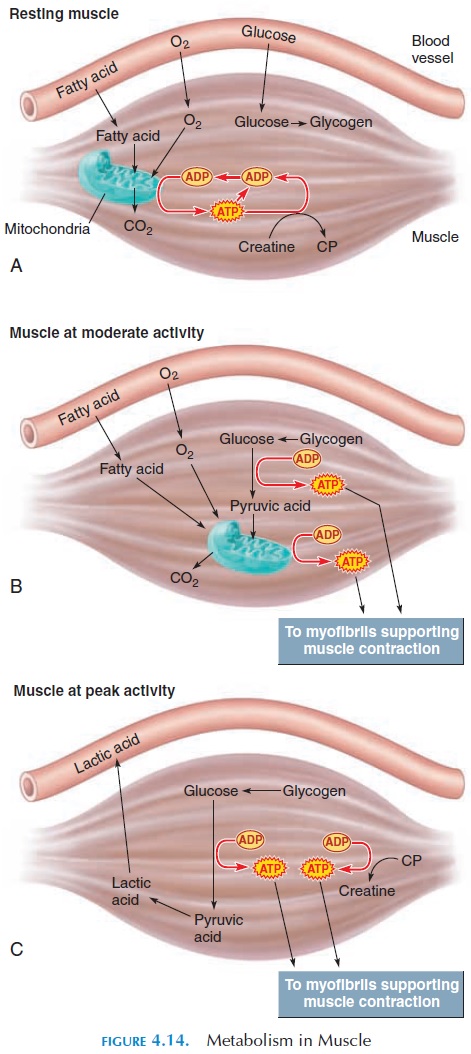
In this type of metabolism, glucose is broken down to pyruvic acid in the cytoplasm of the cell to produce ATP. This process is called glycolysis. Because gly-colysis can take place without the presence of oxy-gen, it is known as anaerobic metabolism. Produc-tion of energy through anaerobic metabolism is an inefficient way to generate ATP. When glucose is bro-ken down to two pyruvic acid molecules, it forms only 2 ATP. However, if the two molecules were used in the TCA cycle in the presence of oxygen in the mi-tochondria, 34 (17 17) ATP could be generated. However, glycolysis is important because it can pro-ceed without the supply of oxygen.
During peak activity, when the muscle is deprived of the ready-made ATP and creatine phosphate, it breaks down glycogen stored in the sarcoplasm to form glucose. This glucose is, in turn, broken down to pyruvic acid and ATP for immediate use. If oxygen is available and adequate, pyruvic acid enters the TCA cycle to produce more ATP.
Glycolysis:
Glucose → 2 pyruvic acid+ 2ATP
If pyruvic acid production by glycolysis is faster than is used by the mitochondria in the TCA cycle, pyruvic acid is converted into lactic acid in the pres-ence of the enzyme lactate dehydrogenase.
Pyruvic acid → lactic acid
Accumulating lactic acid is a disadvantage be-cause it enters the body fluids and easily dissociates into lactate ions and hydrogen ions. This tends to al-ter the pH of the body fluids. Although buffers in the cell and the body fluids try to prevent pH fluctua-tions, their defenses are limited. Eventually, the pH changes slightly and this then alters the function of various enzymes (the activity of enzymes largely de-pends on the pH) and the muscle fiber has difficulty contracting.
In summary, at rest, the demand for ATP is low, and the supply of oxygen is enough for the mito-chondria to produce surplus ATP using fatty acid.
This ATP is used to build up a reserve of creatine phosphate and glycogen from glucose. Fatty acid and glucose are absorbed from the blood.
During moderate levels of activity, the demand for ATP increases. This demand is met by the production of ATP by the mitochondria. Because oxygen supply by the blood is sufficient at this level of activity, the mitochondria form ATP from pyruvic acid. The pyru-vic acid is derived from glucose which, in turn, is de-rived by breaking down glycogen stores. If glycogen stores are depleted, amino acids and lipids may be broken down. Hence, the contribution of glycolysis to the production of energy is minimal.
At high levels of activity, the ATP demands are enor-mous, and the mitochondrial activity is at its maximum. At this point, even if the blood flow is good, the rate of oxygen diffusion from the blood into the cell is not fast enough. The mitochondria can only supply about one-third of the ATP required. The remaining ATP is generated by glycolysis. When the production of pyruvic acid by glycolysis is faster than can be used by the mitochondria, it is converted into lactic acid.
EFFICIENCY OF MUSCLE WORK
The amount of mechanical output from the muscle in relation to the unit of energy put into the muscular sys-tem has been calculated as 20% to 25%. This means that for a given action, the muscle is using four to five times the amount of energy to produce the action. The remaining energy is converted to heat. This is why a lot of heat is produced when muscles are exercised vigor-ously. The heat is dissipated by various regulatory mechanisms in the body, such as production of sweat.
MUSCLE RECOVERY
During the recovery period, the muscle returns to its normal state, and the heat that was produced during metabolism must be dissipated. The muscle reserves of glycogen and creatine phosphate and others must be rebuilt. The lactic acid that was formed must be recycled. It may take several hours for the muscle to recover after a moderate level of activity. After peak levels of activity, it may take a week for the muscle to return to its original state.
Fortunately, the lactic acid produced can be recy-cled; it is converted to pyruvic acid when the level of pyruvic acid is low. This happens soon after exertion. The pyruvic acid made in this way can enter the TCA cycle to produce ATP or it can be converted by special enzymes to glucose and then to glycogen. The lactic acid that enters the blood is taken up by the liver and converted to glucose. The glucose may be stored as glycogen in the liver or it may enter the blood and be used again by skeletal muscle.
During recovery, the oxygen needs of the body rise. This oxygen is used for recovering ATP that was used during muscle contraction. The amount of oxygen required to bring the muscle to its pre-exertion level is known as the oxygen debt. Until the oxygen debt is repaid, the individual continues to breathe at a much faster rate and depth than normal.
The tissue involved in oxygen consumption during the recovery period are the skeletal muscles that must restore ATP, glycogen, and creatine phosphate to for-mer levels. The liver uses ATP to convert lactic acid to glucose. ATP is also used by sweat glands to increase sweat secretion to dissipate heat by evaporation and bring the body temperature back to normal.
MUSCLE FATIGUE
Sometimes, the muscle may find it difficult to con-tract even when stimulated by the nerve. This state is known as muscle fatigue. The cause of fatigue is var- ied and depends on the type of activity. It may be the result of an interruption to the chain of events re-sponsible for muscle contraction—the central ner-vous system (CNS), peripheral nervous system, neu-romuscular junction, and muscle fiber. Following peak activity, the muscle becomes fatigued as a result of the depletion of ATP, creatine phosphate, and glycogen. The lowering of pH (acidity) as a result of lactic acid buildup may also play a part. Prolonged exercise, such as running a marathon, may result in physical damage to the sarcoplasmic reticulum, changes in T tubules, ionic imbalances, and fatigue. Exercise induced alteration in content of CNS neuro-transmitters, such as dopamine, ACh, and serotonin, has been implicated as the cause of psychic or per-ceptual changes that reduce the ability to continue exercising. Fatigue may also be a result of failure of the action potential to cross over the neuromuscular junction. The actual cause of this failure is unknown.
Fatigue may occur more rapidly if the intracellular reserves are low, such as in malnutrition. It is also important to have adequate blood flow to the muscle. Any problems with circulation, such as cardiac prob-lems or tight clothing, that impede blood flow to the muscle, can speed the onset of fatigue. Similarly, any condition that affects the normal blood oxygen con-centration can induce fatigue quickly. Respiratory problems and low levels of oxygen carrying capacity of the blood (e.g., low red blood cell count, reduced hemoglobin) can all result in early fatigue.
MUSCLE PERFORMANCE
The performance of the muscle is measured by the ten-sion or power produced and the duration that a par-ticular activity can be maintained—the endurance. The power and endurance in a muscle are determined by the type of muscle and the level of physical condi-tioning or training.
Types of Muscle Fibers
Skeletal muscle fibers are classified into three types, according to the speed at which they respond to stim-ulus. The three types are fast fibers, slow fibers, andintermediate fibers.
Fast Fibers
The fast fibers are also known as fast twitch, fastglycolytic, or type IIB fibers. Most skeletal musclefibers in the body are fast fibers. These fibers respond to a stimulus in 0.01 second. They are large in diam-eter, with huge reserves of glycogen, densely packed myofibrils, and few mitochondria. The presence of more myofibrils helps these muscles generate a lot of tension; however, because they rely largely on anaer-obic metabolism, they fatigue rapidly. As a result of the lower number of capillaries per unit area, these fibers appear pale to the naked eye.
Slow Fibers
The slow fibers, however, are smaller and take about three times longer to contract after stimulus than fast fibers. Slow fibers are also known as slow twitch or slow oxidative fibers. Slow fibers have an extensivenetwork of capillaries and numerous mitochondria. In addition, slow fibers contain a large amount of myoglobin, a red pigment. Myoglobin is similar to the oxygen carrying hemoglobin protein in the blood. Myoglobin has an affinity for oxygen and makes oxy-gen available when needed. Structurally, these fibers are equipped to contract for a long period without be-coming fatigued (i.e., they have increased endurance). Because these fibers have more blood flowing through them and more myoglobin, these muscles appear red to the naked eye.
Intermediate Fibers
Intermediate fibers have the properties of both slow and fast fibers. They are also known as type IIA or fast oxidative-glycolytic fibers. Similar to fast fibers,they appear pale because they contain less myoglobin. They have more endurance than fast fibers because they have more capillaries per unit area.
The percentage of fast, slow, and intermediate fibers varies. For instance, muscles that must move rapidly for short intervals have a larger proportion of fast fibers—sometimes with no slow fiber at all. The eye muscles and the small muscles of the hand are typical examples. Muscles that are constantly con-tracting to maintain movement and posture, such as calf muscles and back muscles, have a larger propor- tion of slow fiber. The proportion of fast and slow fibers in each muscle is determined genetically. How-ever, it is possible for fibers to change from slow or fast to intermediate type by physical conditioning. For example, if a muscle with more fast fibers is used repeatedly for events that require endurance, the fast fibers may adapt by changing to intermediate fibers.
MUSCLES AND HORMONES
Many hormones affect the metabolism in the muscle fiber. Growth hormone (a hormone secreted by the pi-tuitary gland) together with testosterone (the male hor-mone secreted primarily by the testis) stimulate the formation of contractile proteins and the enlargement of muscles. A synthetic hormone (anabolic steroid) that resembles testosterone is taken by some athletes to increase the size and power of their muscles.
The thyroid hormone can also stimulate the metab-olism of both active and resting muscles.
Related Topics