Chapter: The Massage Connection ANATOMY AND PHYSIOLOGY : Respiratory System
Mechanics of Respiration
Mechanics of Respiration
To fully understand the mechanics of respiration, the learner is encouraged to review the structure of the thoracic cavity.
Respiration refers to the movement of air in andout of the lungs and the exchange of gases. It includes two processes—external and internal respiration.
External respiration refers to all processes involved in the absorption of oxygen and the removal of car-bon dioxide from the air spaces in the lung and pul-monary capillaries. Internal respiration refers to the absorption of oxygen and the removal of carbon diox-ide from tissue cells by the blood surrounding the capillaries. Cellular respiration refers to the process of use of oxygen and production of carbon dioxide for ATP production by cells.
EXTERNAL RESPIRATION
For external respiration, the air must be physically moved from the atmosphere into the lungs. This in-volves increasing and decreasing the size of the tho-racic cavity by movement of the chest wall and action of the respiratory muscles. This process is known as pulmonary ventilation, or breathing. The processof drawing air into the lungs is termed inspiration or inhalation, and the process of moving the airout ofthe lungs is referred to as expiration or exhalation.
The rate at which air flows is not only influenced by the pressure differences between the atmosphere and the thoracic cavity but also by the surface ten-sion in the alveoli, the compliance of the lungs (the ease with which the lungs expand), and the resistance offered by the airways.
Pressure Changes During Pulmonary Ventilation
Knowledge of basic physics will help you understand how air is moved in and out of the thoracic cavity. Ac-cording to Boyle’s law, the pressure inside a closed chamber is inversely related to the volume. Simply, if the volume in a closed container is reduced, the pres-sure of gas in the container increases. For example, if volume is reduced by half, the pressure would dou-ble. If volume is doubled, the pressure would be half of what it was originally. This is the principle behind movement of air in and out of the thoracic cavity.
The parietal pleura lines the inner wall of the tho-racic cavity and the visceral pleura lines the outside of the lungs. The pleural fluid keeps the two membranes in close contact by surface tension. Also, the pressure in the pleural cavity, which is less than that of the at-mosphere, a partial vacuum, keeps the two layers op-posed. When the thoracic cavity increases in volume by the action of muscles and movement of the tho-racic cage, it draws the pleura and, therefore, the lungs with it. This results in a drop in pressure inside the lungs (Remember, when the volume increases, the pressure decreases).Because the nasal cavity commu-nicates with the outside atmosphere, air flows into the lungs to equalize the pressure—inspiration.
Conversely, if the thoracic volume decreases, pres-sure inside the lungs increases (a decrease in volume increases the pressure), and the air flows out of the lungs through the nose to equalize the pressure with that of the atmosphere—expiration. Normal expira-tion is passive, and the change in thoracic volume is caused by the relaxation of the inspiratory muscles and the elastic recoil of the chest wall and the lungs that were stretched during inspiration.
The difference in the oxygen and carbon dioxide levels in the air and blood causes the gases to move by diffusion from the region of higher concentration to that of lower concentration across the alveoli. Thus, carbon dioxide diffuses from the blood to the air and oxygen from air to the blood. Respiration takes place using these physical principles.
Surface Tension and Pulmonary Ventilation
The luminal surface of the alveoli has a layer of fluid. Because water molecules have a greater attraction be-tween each other than with gas molecules, an inward directed force is created. This inward force—surface tension—tries to draw the alveoli into the smallest possible diameter. Therefore, the alveoli tend to col-lapse during expiration. During inspiration, this force opposes entry of air into the alveoli. Surfactant, se-creted by the type II cells, reduces the surface tension and the resistance offered during inspiration.
Lung Compliance
Compliance is the change in volume for a unit changein pressure, and it reflects the ability of the lungs to stretch. If more air volume can be brought into the lung with smaller pressure differences between the lung and the exterior, the lung is considered more com-pliant. For example, lack of surfactant will make the lungs less compliant and make it harder to breathe in. Normal lungs have high compliance because they have elastic tissue that stretch easily. Also, the presence of surfactant reduces surface tension. Respiratory condi-tions that result in scar tissue formation and reduction of elastic fibers, fluid in the lungs, paralysis of respira-tory muscles, or reduced surfactant, lower compliance, increasing the work of breathing. In emphysema, the lungs become more compliant than normal. This, too, is not desirable because there would be a tendency for air to remain in the lungs even after expiration.
Airway Resistance
The resistance to air flow is largely determined by the caliber of the bronchi. By contracting and relaxing the smooth muscle of the bronchi, the resistance to airflow can be modified. When the sympathetic nerves are stimulated, the muscle wall relaxes, increasing the di-ameter of the bronchi and reducing resistance. In con-ditions such as asthma or obstructive pulmonary dis-ease, the airway resistance is increased, making it harder to breathe. Normally, the airways increase in width during inspiration and reduce during expiration.
Presence of mucus and edema in the airways can also affect resistance. In cystic fibrosis, as a result of the presence of a defective gene that carries instruc-tions for a transmembrane protein responsible for the active transport of chloride ions, the transport of salts and water is inefficient. Thick and viscous se-cretions result, with mucous plug formation, inflam-mation, predisposition to infection, and an increase in airway resistance.
RESPIRATION MUSCLES
Aspiratory Muscles
Inspiration is an active process where the muscles of respiration (see Figure 10.10 and Table 1) contract to increase the thoracic volume. The major muscle of in- spiration is the diaphragm. The diaphragm is a dome-shaped, skeletal muscle, projecting into the thoracic cavity. Its origin is along the walls of the thorax and in-sertion is into a centrally placed tendon. The di-aphragm is innervated by the phrenic nerve that arises from cervical segments 3, 4, and 5. When the di-aphragm contracts, the central tendon is pulled down-ward into the abdominal cavity, increasing the diame-ter of the thoracic cavity in the superior-inferior direction. The diaphragm is responsible for 75% of air movement into the lungs.
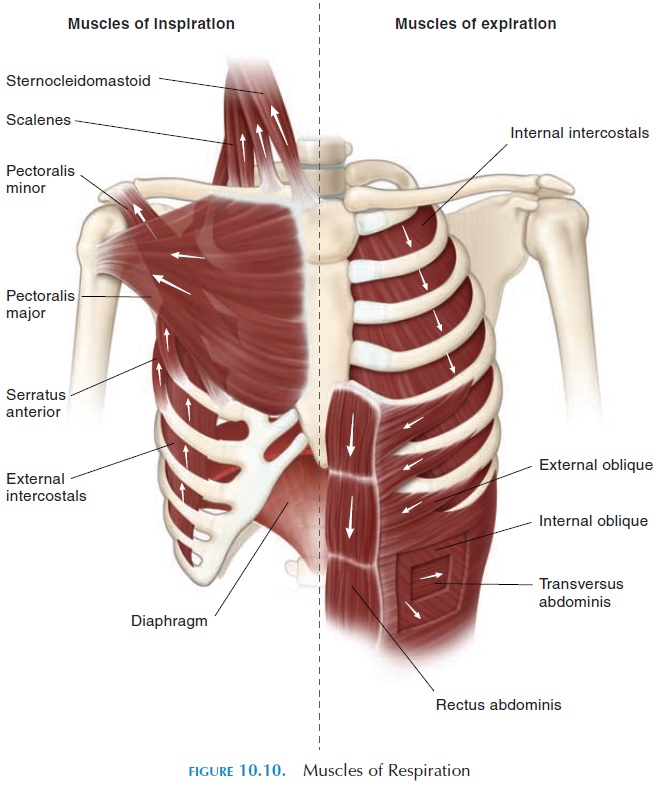
The external intercostal muscles that originate in the upper rib, with insertion in the lower rib, with the fibers running anteriorly and inferiorly, help elevate the ribs. They are innervated by nerves arising from thoracic segments 1 to 12. The ribs articulate with the vertebrae and sternum in a way that the antero-posterior and transverse diameters are increased when they are lifted up by this muscle. The activity of the external intercostals is responsible for bringing approximately 25% of the volume of air into the lungs.
In addition to the diaphragm and external inter-costals, other accessory muscles assist inspiration, in-cluding the sternocleidomastoid, scalenes, serratus anterior, pectoralis minor, pectoralis major, and upper trapezius. The sternocleidomastoid muscles help ele-vate the sternum, and the scalene muscles elevate the upper few ribs, increasing the anteroposterior diameter.
Muscles, such as the serratus anterior, pectoralis major and minor, act as muscles of inspiration by el-evating the ribs or pulling the ribs toward the arms by reverse muscle action (i.e., the insertion is fixed while the origin moves) when the upper limb is fixed in po-sition. The accessory muscles come into play when respiration is forced, as in individuals with asthma.
Expiratory Muscles
During normal respiration, the expiration process is passive and caused by the relaxation of the inspira-tory muscles described above. In forced respiration, expiration becomes active and is assisted by the in-ternal intercostals and transversus thoracis, which help depress the ribs (Figure 10.10 and Table 10.1). The abdominal muscles, which include the external obliques, internal obliques, transversus abdominis, and the rectus abdominis, help compress the ab-domen and force the diaphragm upward.
Other Muscles
The abductor muscles in the larynx contract early in inspiration, pulling the vocal cords apart. The adduc-tor muscles contract reflexively and close the glottis during swallowing or gagging, to prevent food from entering the larynx.
BREATHING PATTERNS, RESPIRATORY VOLUMES, AND CAPACITIES
Breathing Patterns
The number of breaths taken every minute is known as the respiratory rate; a normal adult has a respi-ratory rate of about 12–18 breaths per minute. Chil-dren breathe about 18–20 breaths per minute. The normal rate of inspiration to expiration is 2:1. Eup-nea is the term for normal, quiet breathing. A patternof breathing in which the breathing is shallow, with the chest moving outward, is known as costalbreathing. This movement is a result of the contrac-tion and relaxation of the intercostals muscles that move the ribs. In diaphragmatic breathing, the di-aphragm moves up and down, pushing the abdomi-nal contents outwards.
Respiratory Volumes and Capacities
The respiratory rate and the volume of air moving in and out of the lungs can be easily recorded using the spirometer, or respirometer. The recording ob- tained is thespirogram (see Figure 10.11). An up-ward deflection is produced in the recording when a person breathes in through the mouthpiece of the spirometer. Expiration is recorded as a downward de-flection. The height of deflection reflects the volume breathed; the horizontal axis reflects time.
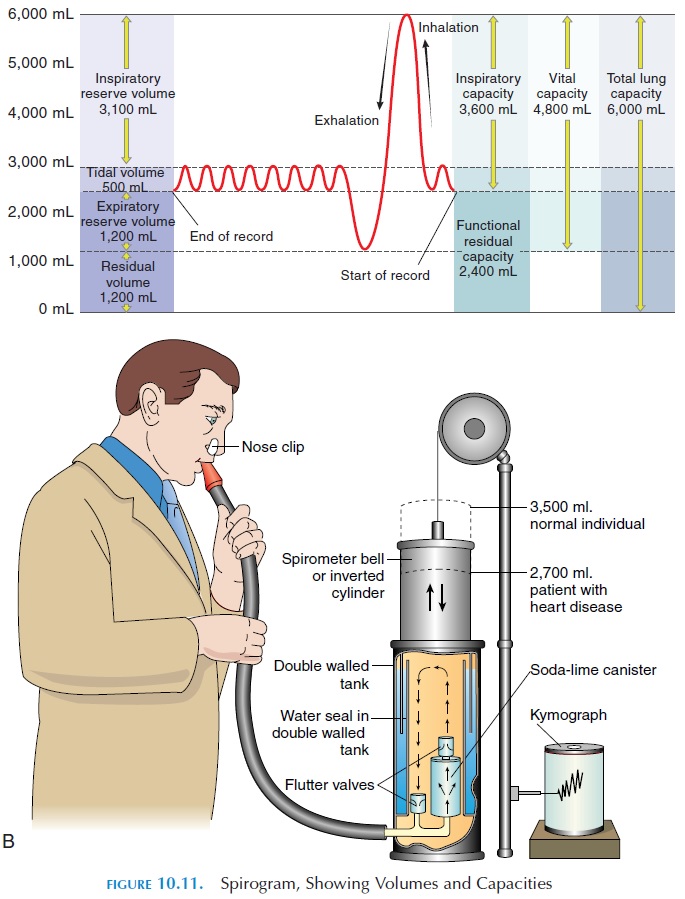
At rest, breathing quietly, an adult takes in about 500 mL (30.5 in3) of air. This is known as tidal vol-ume (see Table 10.2). Thus, the volume of airbreathed in and out every minute during quiet respi-ration is equal to respiratory rate tidal volume and is about 6.0 liters (1.6 gal) per minute. This volume is known as the respiratory minute volume.
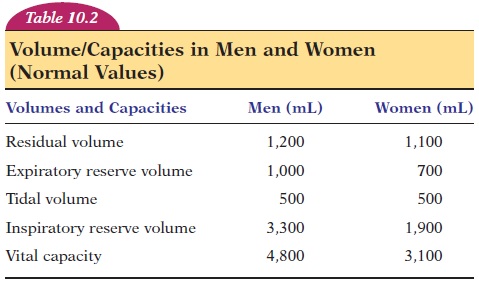
The total volume of air that can be accommodated in the respiratory system is known as the total lungcapacity. This volume averages about 6 liters (1.6gal) in men and 4.2 liters (1.1 gal) in women. Of total lung capacity, the volume of air that a person can breathe in after a normal expiration is much more than the tidal volume. This volume, known as the in-spiratory capacity, includes the inspiratory reserve volume and tidal volume (the approximate values inmen and women are given in Table 10.2). The volume of air that a person can breathe out after a normal ex-piration is known as the expiratory reserve volume.
Even when a person breathes out as forcefully as pos-sible, there is some air remaining in the lungs. This volume of air is known as the residual volume. The residual volume and the expiratory reserve volume together make up the functional residual capacity.Vital capacity is the volume of air that a person canmaximally breathe out after a maximal inspiration. Sometimes, the volume of air a person can forcefully breathe out in the first second is measured. This vol-ume is known as the forced expiratory volume inone second (FEV1), or timed vital capacity. Nor-mally, more than 80% of the total volume expired is expelled in the first second. FEV1 is reduced in con-ditions where obstruction to the airway is present. FEV1 is a useful measurement for monitoring the day-to-day response to drugs, dosage required, and the progress of obstructive diseases in individuals.
A small device known as the peak flow meter is often given to patients to track of their forced expiratory volume. To monitor gas exchange, blood samples may be analyzed to determine the concentration of dissolved gases.
The volumes and capacities, as seen in Table 10.2 differ between men and women. Some volumes also vary with the age and height of the individual. People living in high altitudes and those who have led an ac-tive lifestyle since childhood tend to have larger vital capacities.
The vital capacity of an individual does not matter, what is important is the volume of air that actually reaches the exchange surface—the alveoli. This air, known asalveolar ventilation, is equal to 4.2 liters (1.1 gal) per minute. Because the air that occupies the space in the conducting passages (nasal cavity to the terminal bronchiole) are not involved in gas ex-change, this volume of air is referred to as deadspace air. More specifically, this volume of air isknown as anatomic dead space because this wasted air is a result of the anatomic structure; it occupies about 150 mL (9.2 in3).
Other than the anatomic dead space, some air taken into the lungs may be wasted if the alveoli that they enter do not have a blood supply. Here, although air reaches the exchange surface, there is no blood circulation for gas exchange to take place. The vol-ume of such wasted air is known as physiologicdead space. In normal individuals, this dead space is low. However, in individuals with such conditions as fibrosis of the lungs, the dead space is large and gas exchange is inadequate.
Related Topics