Chapter: Essentials of Anatomy and Physiology: Heart
Histology of the Heart
HISTOLOGY OF THE HEART
Heart Wall
The heart wall is composed of three layers of tissue: the epicar-dium, the myocardium, and the endocardium (figure 12.12). The epicardium (ep-i-kar′dē-ŭm), also called thevisceral pericar-dium, is a thin, serous membrane forming the smooth outer surfaceof the heart. It consists of simple squamous epithelium overlying a layer of loose connective tissue and adipose tissue.
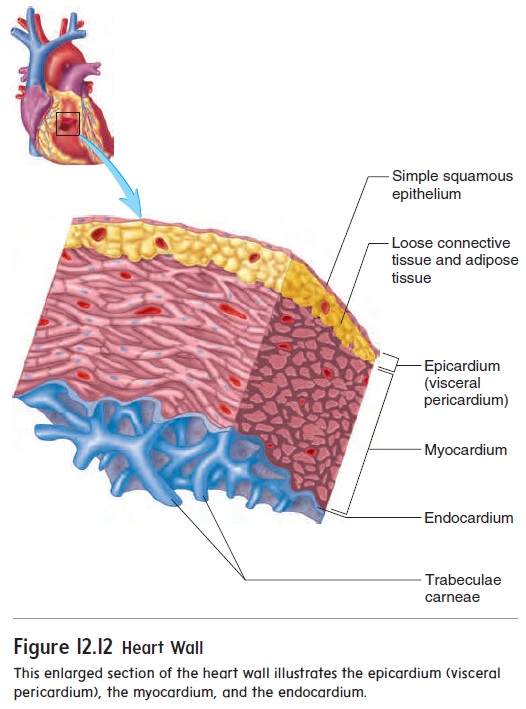
The thick, middle layer of the heart, the myocardium (m -ō -kar′ dē -ŭ m), is composed of cardiac muscle cells and is responsible for contrac-tion of the heart chambers. The smooth inner surface of the heart chambers is the endocardium (en-dō -kar′ dē -ŭ m), which consists of simple squamous epithelium over a layer of connective tissue. The endocardium allows blood to move easily through the heart. The heart valves are formed by folds of endocardium that include a thick layer of connective tissue.
The surfaces of the interior walls of the ventricles are modi-fied by ridges and columns of cardiac muscle called trabeculaecarneae. Smaller muscular ridges are also present in portions ofthe atria.
Cardiac Muscle
Cardiac muscle cells are elongated, branching cells that contain one, or occasionally two, centrally located nuclei (figure 12.13). Cardiac muscle cells contain actin and myosin myofilaments orga-nized to form sarcomeres, which are joined end-to-end to form myofibrils . The actin and myosin myofilaments are responsible for muscle contraction, and their organization gives cardiac muscle a striated (banded) appearance much like that of skeletal muscle. However, the striations are less regularly arranged and less numerous than in skeletal muscle.
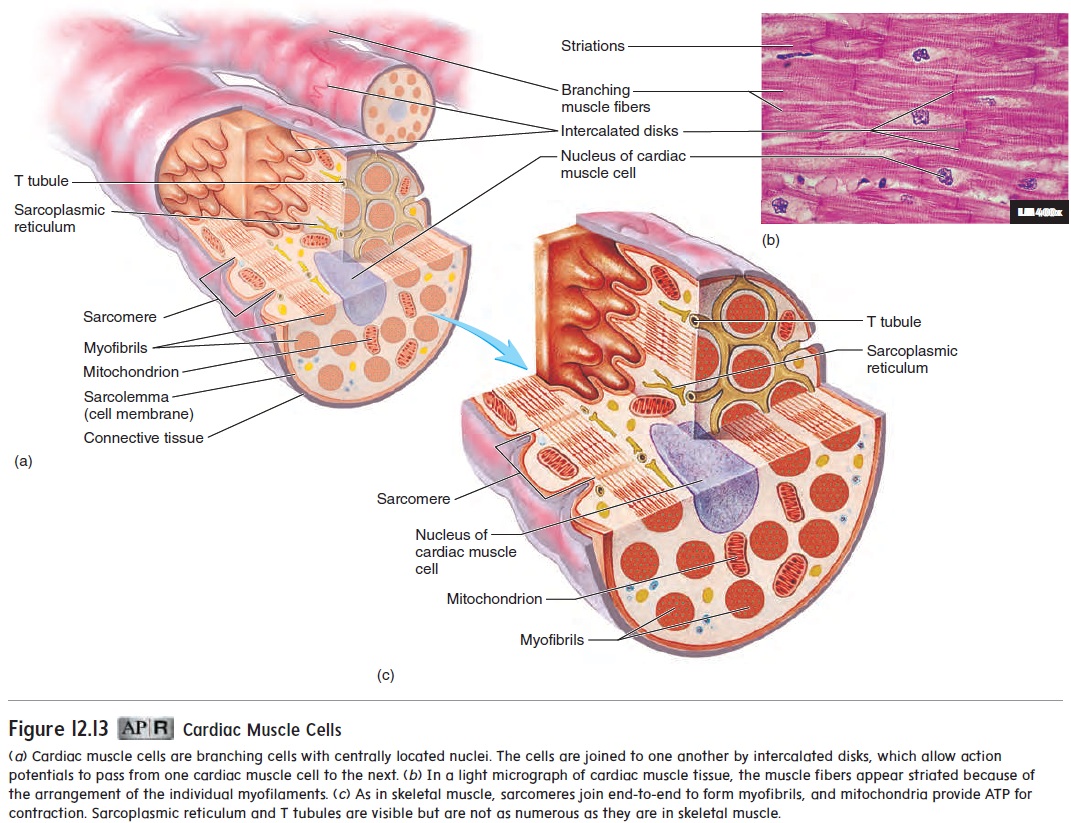
Like skeletal muscle, cardiac muscle relies on Ca2+ and ATP for contraction. Calcium ions enter cardiac muscle cells in response to action potentials and activate the process of contraction much as they do in skeletal muscle. ATP production depends on O2 availability. Cardiac muscle cells are rich in mitochondria, which produce ATP at a rate rapid enough to sustain the normal energy requirements of cardiac muscle. An extensive capillary network provides adequate O2 to the cardiac muscle cells. Unlike skel-etal muscle, cardiac muscle cannot develop a significant oxygen deficit. Development of a large oxygen deficit could result in muscular fatigue and cessation of cardiac muscle contraction.
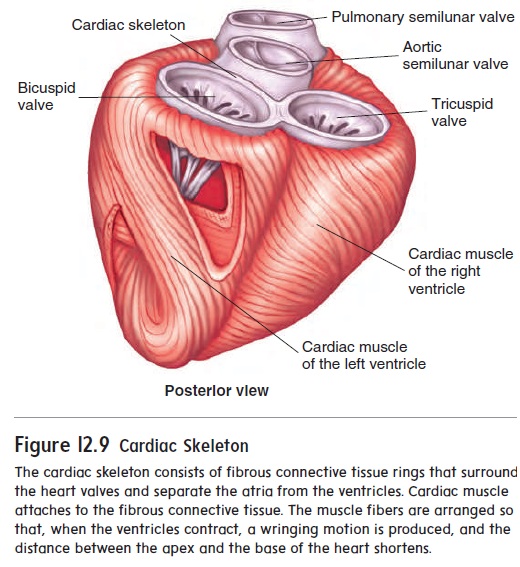
Cardiac muscle cells are organized into spiral bundles or sheets (see figure 12.9). When cardiac muscle fibers contract, not only do the muscle fibers shorten but the spiral bundles twist to compress the contents of the heart chambers. Cardiac muscle cells are bound end-to-end and laterally to adjacent cells by spe-cialized cell-to-cell contacts called intercalated (in-ter′ kă -lā -ted) disks (figure 12.13). The membranes of the intercalated disks arehighly folded, and the adjacent cells fit together, greatly increas-ing contact between them and preventing cells from pulling apart. Specialized cell membrane structures in the intercalated disks called gap junctions allow cytoplasm to flow freely between cells. This enables action potentials to pass quick-ly and easily from one cell to the next. The cardiac muscle cells of the atria or ventricles, therefore, contract at nearly the same time. The heart’s highly coordinated pumping action depends on this characteristic.
Related Topics