Chapter: Human Neuroanatomy(Fundamental and Clinical): Internal Structure of the Spinal Cord
Grey Matter of the Spinal Cord
Grey Matter of the Spinal Cord
Subdivisions of Grey Matter
The grey matter of the spinal cord may be subdivided in more than one manner.
Traditionally, the ventral grey column has been divided into a ventral part, the head, and a dorsal part, the base. Similarly, the dorsal grey column has been subdivided (from anterior to posterior side) into a base, a neck, and a head. These subdivisions have, however, been found to have little importance.
Nuclei in spinal grey matter
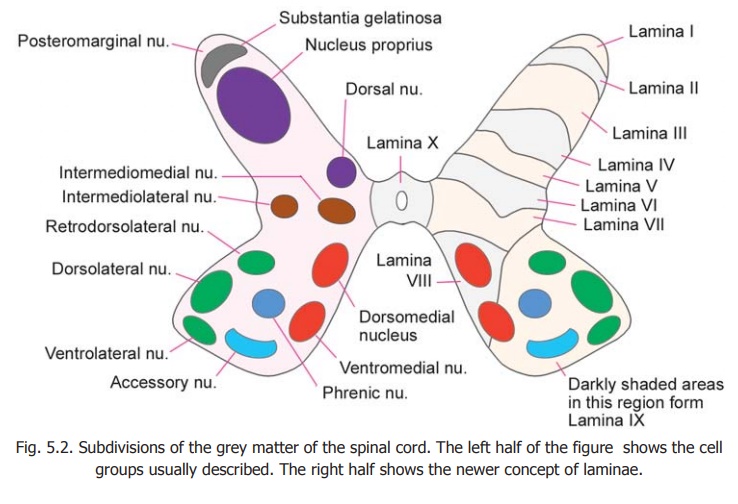
An attempt has been made to recognise discrete collections of neurons (or nuclei) in various regions of the spinal grey matter. These are illustrated in the left half of Fig. 5.2.
The neurons of the ventral grey column (ventral horn cells) are arranged in the form of several discrete and elongated groups. We can recognize
a. medial group which may be subdivided into dorsomedial and ventromedial parts;
b. lateral group consisting of dorsolateral, ventrolateral and retrodorsolateral parts;
c. acentral group represented by the phrenic and accessory nuclei (in the cervical region) and by the lumbosacral nucleus (in the lumbosacral region). Some large cells present along the anterior margin of the ventral grey column are called spinal border cells.
Within the dorsal grey column the following relatively discrete areas may be recognised.
i. The substantia gelatinosa which lies near the apex.
ii. Nucleus proprius (or dorsal funicular group).
iii. Dorsal nucleus (also called the thoracic nucleus or Clark’s column) lying on the medial side of the base.
iv. A thin layer of cells lying superficial to the substantia gelatinosa and constituting the posteromarginal nucleus, also called the marginal zone.
Between the ventral and dorsal grey columns an intermediate zone is sometimes described. This contains the intermediolateraland intermediomedial nuclei (or columns).
Advanced
In the right half of Fig. 5.2.the ventral grey column is shown to be subdivided by a line passing forwards and medially. The part of the column lateral to this line (containing the greater part of the lateral group of nuclei) is present only in the region of the cervical and lumbar enlargements of the spinal cord. From this it can be inferred that the lateral group of nuclei innervate the musculature of the limbs. (Some neurons occupying the position of the ventrolateral group are seen in the first and second sacral segments of the spinal cord. They are believed to supply perineal musculature).
The dorsal nucleus is present only in the thoracic and upper lumbar segments. The intermediolateral group of neurons is present (a) from segments T1 to L2, and (b) in segments S2, S3 and S4.
From the examples given above it will be clear that at a given level of the spinal cord only some of the nuclear groups named in the preceding paragraphs are present.
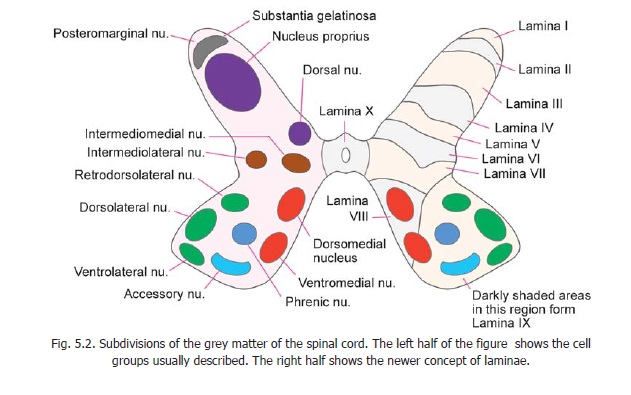
Division of spinal grey matter into laminae
Recent studies have shown that from the point of view of neuronal connections the grey matter of the spinal cord may be divided into ten areas or laminae. These are illustrated in the right half of Fig. 5.2.
Lamina I corresponds to the posteromarginal nucleus (lamina marginalis), lamina II to thesubstantia gelatinosa, and laminae IIIand IV to the nucleus proprius. (Laminae I to IV correspond to the head of the dorsal grey column. Some workers include lamina III in the substantia gelatinosa). Lamina V corresponds to the neck of the dorsal grey column and lamina VI to the base. The lateralpart of lamina V corresponds to the formatio reticularis. Lamina VII corresponds to the intermediate grey matter and includes the intermediomedial, intermediolateral, and dorsal nuclei. It is believed to be made up predominantly of interneurons. At the level of the cervical and lumbar enlargements this lamina extends into the lateral part of the ventral horn. Lamina VIII occupies most of the ventral horn in the thoracic segments, but at the level of the limb enlargements it is confined to the medial part of the ventral horn.Lamina IX is made up of the various discrete groups of ventral horn cells already described. Lamina X forms the grey matter around the central canal.
From the point of view of function, and of neuronal organisation, it has been proposed that (instead of division into ventral and dorsal grey columns) the spinal grey matter should be divided into a central core where the organisation of neurons is diffuse and non-discriminative; and into dorsal and ventral appendages. Laminae VII and VIII have been (tentatively) assigned to the centralcore, laminae I to VI to the dorsal appendage, and lamina IX to the ventral appendage.
Significance of Neurons in Grey Matter of Spinal Cord
The ventral horn cells of the spinal cord may be functionally divided into three major categories as follows.
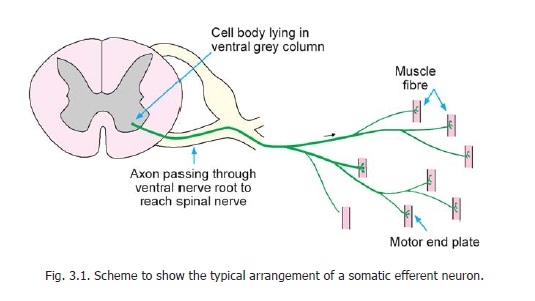
a. The most prominent neurons with large cell bodies and prominent Nissl substance are designated alpha neurons. These are somatic efferent neurons (Fig. 3.1). Their axons (alpha efferents) leave the spinal cord through the ventral nerve roots of spinal nerves and innervate skeletal muscle. They occupy lamina IX of the ventral grey column.
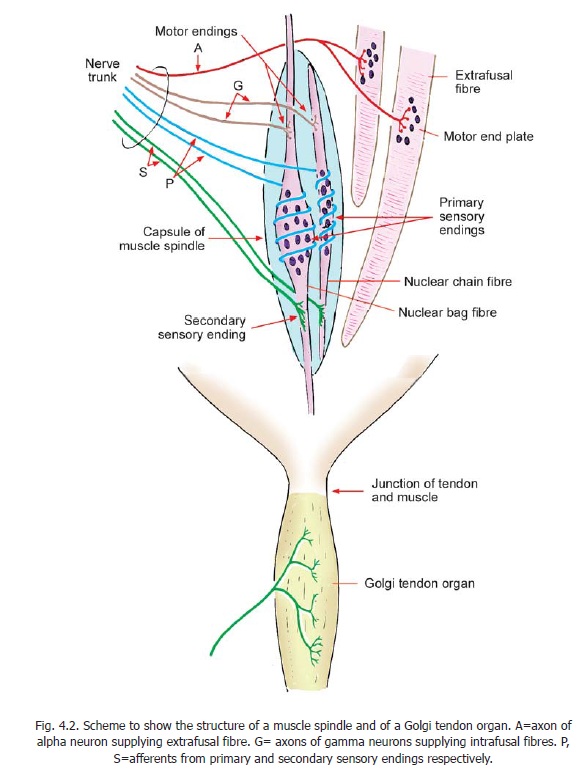
b. Some smaller neurons designated as gamma neurons are also located in lamina IX. They supply intrafusal fibres of muscle spindles (Fig. 4.2). Sensory impulses arising in the spindle travel to the spinal cord and reach the alpha neurons. The gamma neurons thus influence the activity of alpha neurons indirectly through muscle spindles.
c. A considerable number of smaller neurons in the ventral grey column are internuncial neurons. They are most abundant in Lamina VII. Some ramifications of the central processes of cells in the dorsal nerve root ganglia (bringing afferent impulses from the periphery), and axons descending from higher centres, terminate in relation to these internuncial neurons. The axons of internuncial neurons convey these impulses to alpha and gamma neurons.
d. Another variety of neuron that is believed (on physiological grounds) to exist in the ventral grey column is the so calledRenshaw cell. These cells receive the terminations of collaterals arising from the axons of alpha neurons. The axons of Renshaw cells carry the impulses back to the cell bodies of the same alpha neurons, and thus help to regulate their activity.
The neurons of the dorsal grey column may be subdivided as follows.
a. Some of these are internuncial neurons similar to those in the ventral grey column.
b. Many dorsal column neurons receive afferent impulses through the central processes of neurons in dorsal nerve root ganglia. These dorsal column neurons give off axons that enter the white matter of the spinal cord either on the same or opposite side.
These axons may behave in one of the following ways.
1. They may ascend or descend for some segments before terminating in relation to neurons at other levels of the spinal cord. Such axons constitute intersegmental tracts.
2. A considerable number of axons arising from dorsal column neurons run upwards in the spinal cord and constitute ascending tracts which terminate in various masses of grey matter in the brain. These tracts form a considerable part of the white matter of the spinal cord.
The neurons of the intermediolateral group (lateral grey column) are visceral efferent neurons. They are present at two levels of the spinal cord.
a. One group is present in the thoracic and upper two or three lumbar segments. These are preganglionic neurons of the sympathetic nervous system. Their axons terminate in relation to postganglionic neurons in sympathetic ganglia (and occasionally in some other situations). Axons of these postganglionic neurons are distributed to various organs, and to blood vessels (Figs. 19.1, 19.2).
b. The second group of visceral efferent neurons is found in the second, third and fourth sacral segments of the spinal cord. These are preganglionic neurons of the parasympathetic nervous system. Their axons leave the spinal cord through the ventral nerve roots to reach spinal nerves. They leave the spinal nerves as the pelvic splanchnic nerves which are distributed to some viscera in the pelvis and abdomen (Fig. 19.6). They end by synapsing with ganglion cells located in intimate relationship to the viscera concerned. The postganglionic fibres arising in these ganglia are short and supply smooth muscle and glands in these viscera.
Advanced
The location of the various types of neurons described above in relation to laminae of spinal grey matter is of interest.
Afferent fibres carrying sensations from the skin end predominantly in laminae I to IV. Proprioceptive impulses reach laminae V and VI. These laminae also receive numerous fibres from the cerebral cortex.
Lamina VII gives off fibres that reach the midbrain and cerebellum (through spino-cerebellar, spino-tectal and spino-reticular tracts). It receives fibres from these regions through tectospinal, reticulo-spinal and rubrospinal tracts.
Renshaw cells are located in a forward extension of lamina VII (into the interval between laminae VIII and IX).
Lamina VIII is made up mainly of interneurons that receive fibres from various sources. They give efferents to gamma neurons and thus influence muscle spindles.
Lamina IX contains the alpha and gamma neurons that give off efferent fibres to skeletal muscle. Motor neurons in different parts of the ventral grey column show remarkable differences in the orientation and extent of their dendritic fields. Many of the dendrites of neurons in the ventromedial, central and ventrolateral columns run longitudinally in the form of bundles. These neurons, therefore, come into intimate contact with each other. This arrangement is seen in relation to neurons that supply postural muscles. In contrast, the dendrites of neurons in the dorsolateral column have very little contact with those of neighbouring neurons. This column supplies muscles in distal parts of the limbs and the discrete nature of the neurons may be associated with fine control necessary for movements produced by these muscles.
Related Topics