Chapter: Essentials of Anatomy and Physiology: Development, Heredity, and aging
Genes
Genes
Each chromosome contains thousands of genes. Each gene con-sists of a certain portion of a DNA molecule, but not necessarily a continuous stretch of DNA. Genes determine the proteins in a cell.
Both chromosomes of a given pair contain similar but not necessarily identical genes. Similar genes on homologous chro-mosomes are called alleles (ă-lēlz′ ). If the two allelic genes are identical, the person is homozygous (hō-mō-zı̄′ gŭs) for the trait specified by that gene. If the two alleles are slightly different, the person is heterozygous (het′ er-ō-zı̄′ gŭs) for the trait. All the genes in one homologous set of 23 chromosomes in one individual con-stitute that person’s genome (jē′ nōm).
Through the processes of meiosis, gamete formation, and fertilization, the distribution of genes received from each parent is essentially random. This random distribution is influenced by sev-eral factors, however. First, all the genes on a given chromosome are linked; that is, they tend to be inherited as a set rather than as individual genes because chromosomes, not individual genes, segregate during meiosis. Second, homologous chromosomes may exchange genetic information during meiosis by crossing over.
Segregation errors may occur during meiosis. As the chro-mosomes separate, the two members of a homologous pair may not segregate. As a result, one of the daughter cells receives both chromosomes of a given pair, and the other daughter cell receives none. When the gametes are fertilized, the resulting zygote has either 47 chromosomes or 45 chromosomes, rather than the normal 46. When this condition results in an abnormal autosomal chromo-some number, it is usually, but not always, lethal and is one reason for a high rate of early embryo loss. Down syndrome, or trisomy 21, which results when there are three #21 chromosomes, is one of the few autosomal trisomies that is not lethal. In contrast, sex chromo-some abnormalities are not usually lethal. For example, TurnerSyndrome ormonosomy X,in which only one X chromosome ispresent, results in sterility and abnormal sexual development but does not affect the mental development of the female.
Dominant and Recessive Genes
Most human genetic traits are recognized because defective alleles for those traits exist in the population. For example, on chromo-some 11 is a gene that produces an enzyme necessary for the synthesis of melanin, the pigment responsible for skin, hair, and eye color . An abnormal allele, however, produces a defective enzyme not capable of catalyzing one of the steps in melanin synthesis. If a given person inherits two defective alleles, a homozygous condition, the person is unable to produce melanin and therefore lacks normal pigment. This condition is referred to as albinism (al′ bi-nizm).
For many genetic traits, the effects of one allele for that trait can mask the effect of another allele for the same trait. For example, a person who is heterozygous for the melanin-producing enzyme gene has one normal gene for melanin production and one defective gene for melanin production. One copy of the gene and its resulting enzymes are enough to make normal melanin. As a result, the person who is heterozygous produces melanin and appears normal. In this case, the allele that produces the normal enzyme is said to be dominant, whereas the allele producing the abnormal enzyme is recessive. By convention, dominant traits are indicated by uppercase letters, and recessive traits are indicated by lowercase letters. In this example, the letter A designates the domi-nant normal, pigmented allele, the letter a the recessive abnormal allele. Not all dominant traits are the normal condition, and not all recessive traits are abnormal. Many examples exist of abnormaldominant traits. For example, Huntington disease, a degenerative neurological disease, is caused by an abnormal dominant gene.
The possible combinations of dominant and recessive alleles for normal melanin production versus albinism are AA (homo-zygous dominant, normal), Aa (heterozygous, normal), and aa (homozygous recessive, albino). The combination of alleles a person has for a given trait is called the genotype (jen′ ō-tı̄p). The person’s appearance is called the phenotype (fē′ nō-tı̄p). A person with the genotype AA or Aa has the phenotype of normal pigmen-tation, whereas a person with the genotype aa has the phenotype of albinism. The recessive trait is expressed only when no allele for the dominant trait is present. A carrier is a heterozygous person with an abnormal recessive gene but a normal phenotype because the normal dominant allele for that gene is also present.
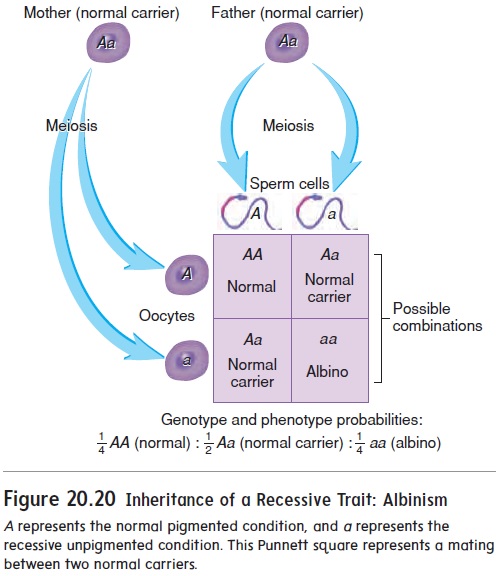
The inheritance of dominant and recessive traits can be deter-mined if the genotypes of the parents are known. For example, if an albino person (aa) mates with a heterozygous normal person (Aa), the probability that the child will be albino (aa) is one-half, and the probability that the child will be normal (Aa) is one-half. If two carriers (Aa) mate, the probability that the child will be homozygous dominant (AA) or homozygous recessive (aa) is one-fourth for either genotype. The probability that the child will be heterozygous (Aa) is one-half. Such a probability can be determined easily by using a table called a Punnett square (figure 20.20).
Sex-Linked Traits
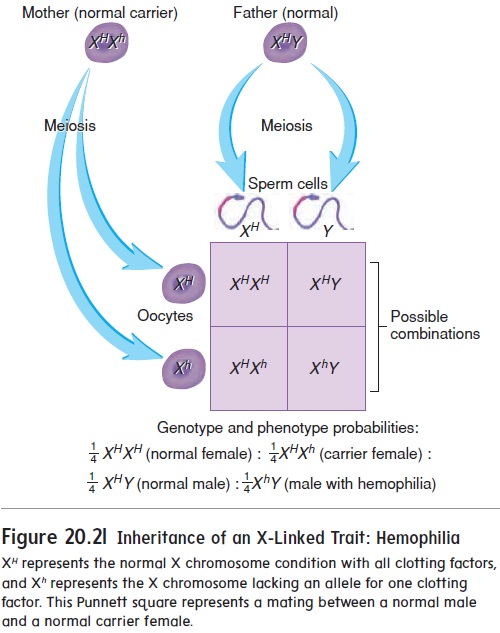
Traits affected by genes on the sex chromosomes are called sex-linked traits. Most sex-linked traits are X-linked; that is,they are on the X chromosome. Only a few Y-linked traits exist, largely because the Y chromosome is very small. An example of an X-linked trait is hemophilia A (classic hemophilia), in which the person is unable to produce one of the clotting factors . Consequently, clotting is impaired, and persistent bleeding can occur, either spontaneously or as a result of an injury. Hemophilia A is a recessive trait, and the allele for the trait is located on the X chromosome. The possible genotypes and phe-notypes are therefore
XHXH (normal homozygous female)
XHXh (normal heterozygous female)
XhXh (hemophiliac homozygous female)
XHY (normal male)
XhY (hemophiliac male)
Other Types of Gene Expression
In some cases, the dominant allele does not completely mask the effects of the recessive allele. This is called incomplete domi-nance. An example of incomplete dominance is sickle-cell anemia, in which the hemoglobin produced by the gene is abnormal. The result is sickle-shaped red blood cells, which are likely to stick in capillaries and tend to rupture more easily than normal red blood cells. The normal hemoglobin allele (S) is dominant over the sickle-cell allele (s). A normal person (SS) has normal hemo-globin, and a person with sickle-cell anemia (ss) has abnormal hemoglobin. A person who is heterozygous (Ss) has half normal hemoglobin and half abnormal hemoglobin and usually has only a few sickle-shaped red blood cells. This condition is called sickle-cell trait.
In another type of gene expression, called codominance (kō-dom′ i-năns), two alleles can combine to produce an effect without either of them being dominant or recessive. For example, a person with type AB blood has A antigens and B antigens on the surface of his or her red blood cells . The antigens result from a gene that causes the production of the A antigen and a different gene that causes the production of the B antigen, and neither gene is dominant or recessive in relation to the other.
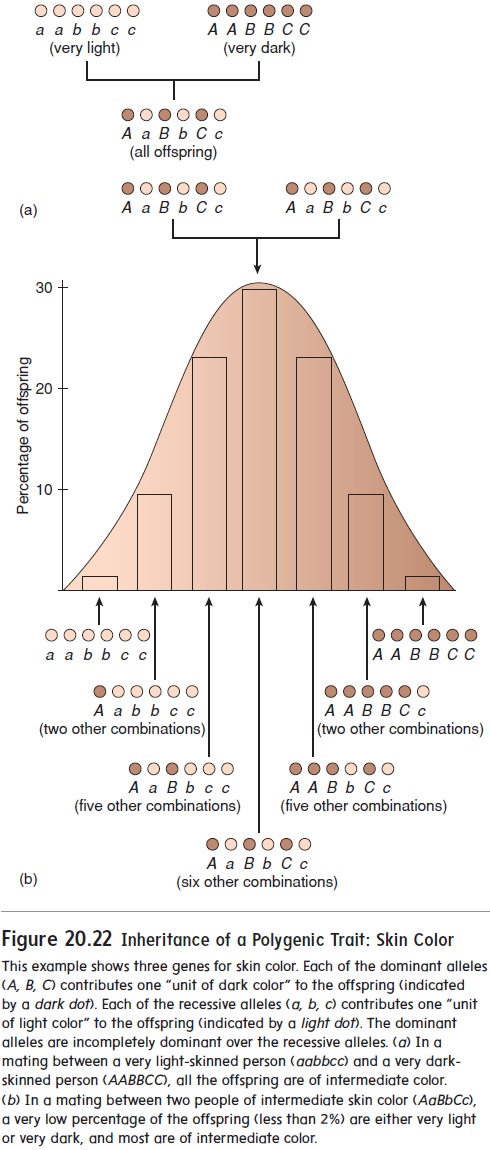
Many traits, called polygenic (pol-ē-jen′ ik) traits, are deter-mined by the expression of multiple genes on different chromo-somes. Examples are height, intelligence, eye color, and skin color. Polygenic traits typically have a great amount of variability. For example, there are many shades of eye color and skin color (figure 20.22).
Related Topics