Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Sensory systems
Electrical communication - Fishes
Electrical communication
Most fishes that produce electricity use it for communication. Signals are species specific and certain aspects of theEOD, such as amplitude, frequency, and pulse length, can be modified to exchange information about species, sex,size, maturation state, location, distance, and probably individual identification (von der Emde 1998; Collin &Whitehead 2004).
Agonistic interactions involving frequency shifts play an important role in dominance interactions in many electric fishes. South American gymnotiform knife fishes have individual characteristic waveforms to their EODs. In Gymnotuscarapo (Gymnotidae), rapid increases and decreases in frequency indicate threat, whereas submissive individuals cease discharging (Black-Cleworth 1970). Within this species, individuals with higher EOD frequencies are consistently dominant. Male Eigenmannia virescens(Sternopygidae)will mouth fight until a dominance hierarchy is established, the ultimate dominant male assuming the lowest discharge frequency. Females compete for spawning territories, and dominant females have the highest frequency(Hopkins 1972, 1974a, 1974b). In the genus Sternopygus(Sternopygidae), mature males discharge at about an octave below the discharge of mature females, which is 120–240 Hz (Feng 1991). The Brown Ghost Knife fish(Apteronotus leptorhynchus) also demonstrates a variety of EODs that convey different meanings, including genderand social status (Zakon et al. 2002).
The diverse mormyriform elephant fishes (Mormyridae)of Africa use EODs for orientation, territorial interactions, species recognition, individual recognition, courtship, and to communicate social status (Carlson 2002; Terleph &Møller 2003). Mormyrids receive information based both on the waveform of the EOD and on intervals between discharges. Variations in discharge interval of fractions ofa millisecond are detectable by the fish. EODs are again both species and sex specific among different life history stages. Males typically have a two to three times longer pulse duration than females. Interactions include cessation and frequency modulation of EODs (“bursts”, “buzzes”, and “rasps”), echoing, and dueting. Males alternate their outputs with other males, whereas females synchronize their outputs with investigating males. A male can determine the sex of a conspecific by “listening” to the response to his electric pulses. In direct analogy to gymnotiformbehavior, the mormyriform Gymnarchus niloticus ceases discharging just prior to an attack on a conspecific, uses bursts of discharge pulses when aggressive, and modulates its frequency by 1–30 Hz as a submissive gesture (Møller1980; Hopkins 1986; Bleckmann 1993).
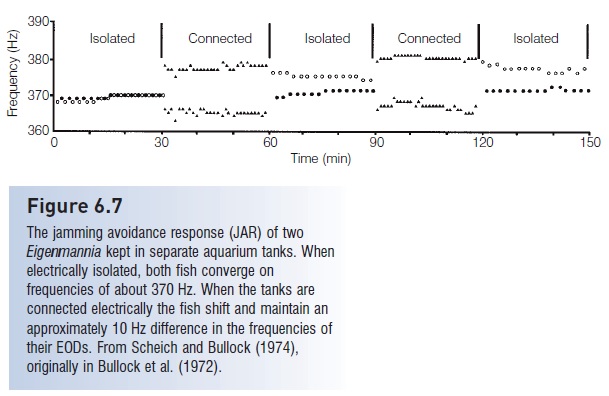
Figure 6.7
The jamming avoidance response (JAR) of twoEigenmannia kept in separate aquarium tanks. When electrically isolated, both fish converge on frequencies of about 370 Hz. When the tanks are connected electrically the fish shift and maintain an approximately 10 Hz difference in the frequencies of their EODs. From Scheich and Bullock (1974),originally in Bullock et al. (1972).
Agonistic interactions include interference with a conspecific c’s electroreception. In Gymnotus carapo, dominant fish often shift their discharges to coincide with the short interval when a subordinate would be analyzing its own output, which could impair the subordinate’s ability toelectro locate. Such interference is overcome in gymnotiformssuch asEigenmannia by a jamming avoidance response(JAR), in which fish shift their EOD frequencies when they get near one another, thereby preventing interference with one another’s ability to electro locate. Fish in a social group maintain a 10–15 Hz difference with their neighbors so that each individual has a “personal” discharge frequency (see Feng 1991). When several Eigenmannia were kept in separate tanks and all the tanks were connected by electrical wires, the fish shifted their frequencies to an average separation of 7 Hz (Fig. 6.7). The Brown Ghost Knife fishalso demonstrates a JAR (Zakon et al. 2002), and, in several other species, fish in a social group have non-overlapping frequencies (Bullock et al. 1972; Hagedorn 1986).
This discussion of electrical communication would be incomplete without some consideration of the source of the electrical signals – the electric organs. The electric-generating cells of electric organs are referred to as electrocytes,and often are dislike modified muscle cells, called electroplaques. When stimulated, ion flux across the cell membranes creates a small electric current, and because the cells are arranged in a column and discharge simultaneously, they produce an additive effect. A sizeable stack of cells can produce a considerable current – like many small batteries connected in series (Feng 1991). Although electrocytes of most electric fishes are modified muscle cells, South American electric fishes of the family Apteronotidae utilize modified neurons (Zupanc 2002).
The generation and detection of weak electric fields is particularly well developed in several groups of freshwater tropical fishes living in murky waters with poor visibility, such as the Gymnotiformes of South America and the Mormyridae of Africa. The EOD of some species are brief pulses released at irregular intervals, whereas other species continuously produce oscillating, high-frequency waves of electricity (Zakon et al. 2002). The resulting electric field surrounds the fish (see Fig. 6.6) and any changes in the field are detected by the fish’s tuberous organs. Bending the body would distort the electric field, so these fishes typically rely on their extensive dorsal or anal fins for propulsion so that they can maintain a straight body posture.
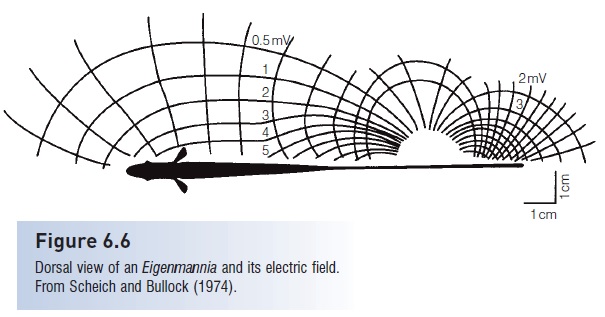
Figure 6.6
Dorsal view of an Eigenmannia and its electric field.From Scheich and Bullock (1974).
The production of weak electric fields, as demonstrated in the gymnotids and mormyrids, requires considerable coordination by the central nervous system. In the South American gymnotid Apteronotus, the electric organs are controlled by pacemaker cells in the medulla, which are regulated by input from two clusters of neurons elsewhere in the brain (Zakon et al. 2002). The location and function of the pacemaker neurons of the African mormyrids is somewhat similar – a remarkable coincidence considering the two groups are believed to have evolved their EODcapabilities independently (Carlson 2002).
The African mochokid catfishes also are believed to produce and detect weak electric fields for object detection or communication (Hagedorn et al. 1990). The electric organ is located dorsally on these catfishes and has apparently evolved from one of the muscles associated with sound production, which occurs by stridulation of the pectoral spines.
Related Topics