Chapter: The Diversity of Fishes: Biology, Evolution, and Ecology: Sensory systems
Chemoreception - Fishes
Chemoreception
The aquatic environment is filled with a wide variety of chemical signals because so many chemicals dissolve in water. Therefore, fishes can learn a great deal about their environment through chemoreception, which often is used foring and identifying food, locating habitat, detecting and avoiding predators, and communicating with conspecifics. The sense of smell (olfaction) helps fishes detect abroad range of chemical stimuli, whereas the sense of taste(gustation) is primarily focused on food recognition(Sorensen & Caprio 1998).
Smell
The organs of smell in fishes are contained within olfactory chambers (Sorensen & Caprio 1998). Jawed fishes have paired olfactory chambers, each of which has an incurrent and excurrent nostril. Cilia within the chambers move water into and out of the nostrils, which usually are small pits separated by a flap of skin, but may be tubular in some fishes such as the Bowfin (Amiidae) and eels(Anguillidae). Hagfishes and lampreys have only a singlenaris and a median olfactory sac nostril. In hagfishes anasohypophyseal duct connects with the pharynx so that hagfishes can smell water as it moves to the gills. In lampreys, however, the lone medial nostril leads to an olfactorychamber in a dead-end nasopharyngeal pouch. Interests the olfactory chambers also are dead-end sacs that do not lead to the pharynx, except in a few cases such as stargazers (Uranoscopidae). The nares of elasmobranchs are located ventrally on the snout and also are not connected to the pharynx. Chimaeras (Holocephali) and lungfishes (Ceratodontiformes) have paired nares that connect to the oral cavity.
Each olfactory sac is lined with a highly folded olfactory epithelium, often arranged in rosettes (Fig. 6.9). Molecules of odorants bind to receptor proteins on membranes of the receptor cells in the sensory epithelium. The receptors cells then send nerve impulses to the brain (Hara 1993). Structure of the rosettes and olfactory sacs is related to the olfactory sensitivity of a fish. The more extensive the lamellar folding, the greater the surface area available for sensory cells and the more sensitive the sense of smell.Freshwater eels (Anguilla) are known for their extremely keen sense of smell and have from 69 to 93 folds in each rosette. Perch(Perca), with less sensitive olfactory capabilities, have 13–18folds in each rosette.
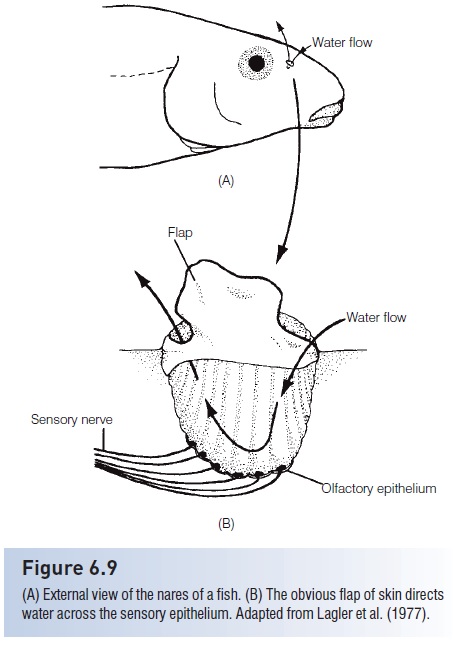
Figure 6.9
(A) External view of the nares of a fish. (B) The obvious flap of skin directs water across the sensory epithelium. Adapted from Lagler et al. (1977).
Fishes are extremely sensitive to certain types of chemicals. Amino acids, particularly those of fairly simple structureand with certain attached groups, are detectable by many fishes at concentrations of around 10–10 mol/L (seeHara 1993). Other compounds that are detectable by some fishes at very low concentrations include bile acids(10–9mol/L), salmon gonadotropin-releasing hormone(10–15 mol/L), and some sex steroids (10–12 mol/L). This ability to detect such small concentrations of certain chemicals makes olfaction valuable in homing in salmon (Stewartet al. 2004; Mechanisms of migration), andin habitat location for some other fishes.
Sea Lamprey (Petromyzon marinus) also areanadromous, and although they do not return to their home streams, they rely on olfaction to identify a suitable spawning stream. A chemical signal released by juveniles (ammocoetes)provides a signal to adults that the stream apparently provides a suitable spawning and nursery habitat, and sexually mature males release another pheromone that attracts mature females (Wagner et al. 2006). This knowledge is being used to try to control Sea Lamprey, which parasitize larger fish, by diverting them during their spawning migration(see Petromyzontiforms).
Juvenile eels in New Zealand may use odor to locate suitable habitats as they migrate upstream after hatching atsea. Glass eels of both the Longfin Eel (Anguilla dieffenbachii)and Shortfin Eel (A. australis) preferred water from their river of capture over well water, and the Shortfin Eels preferred water from lowland streams, where they tend to occur, over water from the mainstream of a river (McCleave& Jellyman 2002).
Olfactory cues also can be used to locate mates, and some fishes exhibit different olfactory sensitivities between sexes. In deep sea ceratioid anglerfishes, males have enlarged olfactory organs, olfactory nerves, and olfactory lobes in the brain, whereas these features are much smaller among females. In these species, females, which are much larger, are thought to release species-specific pheromones that the smaller, more mobile males use to locate them. Males then attach themselves to the females and spend the rest of their lives as parasitic sperm factories (see Thedeep sea). Gilthead Seabream (Sparus aurata) are sensitive to the excreted body fluids of sexually mature conspecifics(Hubbard et al. 2003), and male Brown Trout (Salmotrutta) and Lake Whitefish (Coreg onus clupeaformis) both show courtship behavior when exposed to a prostaglandin released by females ready to spawn (Laberge & Hara 2003).
Olfaction may also be used to detect and avoid predators. Juvenile Lemon Sharks (Negaprion brevirostris) react to the odor of organic compounds produced by American crocodiles (Crocodylus acutus) that prey on small sharks where they co-occur (Hueter et al. 2004). Many fishes respond to chemical alarm cues released from injured conspecifics, or other prey species with which they occur(Brown 2003). The alarm substance, or a metabolite of it, also is present in the feces of predatory species and would therefore also be present in the water nearby. This allows potential prey to inspect predators and assess their potential threat(see Shoaling and search).
Chemical contaminants can interfere with olfaction and thereby disrupt important interspecific communication. Forexample, cadmium accumulates on the olfactory epithelium of Rainbow Trout (Oncorhynchus mykiss) and affects fish social behavior, including blocking their ability to detectalarm substance (Scott et al. 2003; Sloman et al. 2003).
Taste
The sense of taste is used primarily for food recognition. The chemosensory cells responsible for taste are located in and around the mouth, including barbels and lips, and may also be found on the fins and trunk (Sorensen & Caprio1998). Taste receptor cells are often clustered into taste buds, which can contain 30–100 sensory cells, or they may occur individually on parts of some fishes. These solitary chemosensory cells can be numerous, with up to 4000per mm2 in some minnows (Cyprinidae). Sensory neurons synapse with the sensory cells at their basal surface, and when stimulus molecules bind to receptors on the sensory cells, neurotransmitters are released that affect the generation of action potentials by the sensory neuron that carries the signals to the gustatory centers of the brain (Sorensen & Caprio 1998). Toxins, amino acids, and bile salts can stimulate taste receptors at sensitivity thresholds similar to those of olfactory receptors (see Hara 1993).
Related Topics