Chapter: Essentials of Psychiatry: Cognitive Neuroscience and Neuropsychology
Cellular Basis of Memory
Cellular Basis of Memory
On the basis of lesion analytical studies, the
hippocampus and related structures have been implicated as a site for the
con-solidation of memory. The question naturally arises out of the properties
of the neurons in this area that might permit the as-sociation of information.
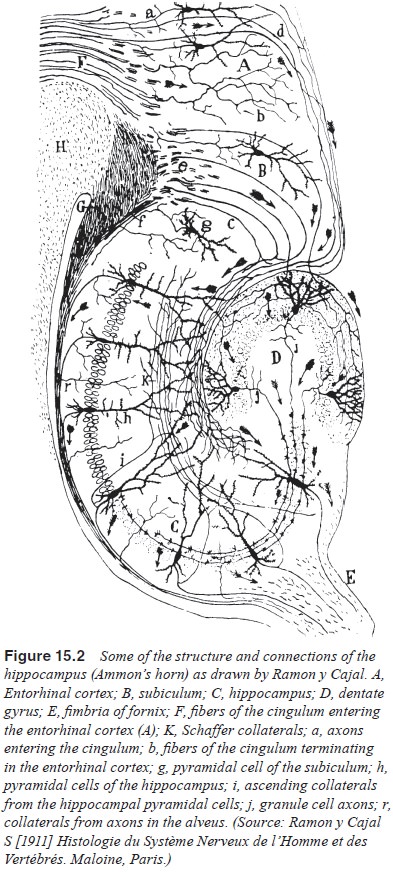
Neuroanatomically (Figure 15.2), the hippocampal formation consists of the
fields of Ammon’s horn (regions CA1, CA2, CA3),
the dentate gyrus and the subiculum. Afferent pathways from the entorhinal
cortex project to the hip-pocampus via the perforant pathway and synapse on the
granular cells in the dentate gyrus. The entorhinal cortex itself receives
cortical inputs from polysensory associational regions in the frontal, temporal
and parietal lobes. Within the hippocampal formation, the granule cells in
region CA3 also project to the CA1 region through the fimbria
fornix, which also projects to the subiculum. The subiculum is the major
efferent pathway, project-ing to a number of cortical regions but also
projecting back to the entorhinal cortex, completing the loop.
For learning and memory to occur, there must be
plastic changes such that the structure and functional characteristics of nerve
cells and their interconnections are altered. Much of the re-search into the
processes of synaptic plasticity underlying learn-ing and memory has been
conducted in invertebrates (Kandel et al.,
1983; Kandel, 1991) through examination of reflexive learning processes (habituation, sensitization and classical
conditioning).
Regarding the mammalian brain, Bliss and Lomo
(1973) were the first to demonstrate that repeated stimulation of the
afferent pathways to the dentate granule cells of
the hippocampus of the rabbit produced an excitatory potential in the
postsynaptic hippocampal neurons lasting for hours. Recording in intact
ani-mals has shown potentials that lasted for days and weeks. They termed this
increased facilitation as a result of repeated stimula-tion long-term
potentiation (LTP).
With respect to area CA1 of the
hippocampus, studies have shown that LTP occurs only when a number of input
pathways have been stimulated. This is known as the criterion of
coopera-tivity. When distinct weak and strong excitatory inputs impinge on a
pyramidal nerve cell, the weak input becomes potentiated through association
with a strong input. This is known as the cri-terion of associativity. Finally,
the criterion of specificity refers to
the fact that strong repeated stimulation in one
synaptic pathway is specific only to that stimulated pathway. Unstimulated
syn-apses on the same cell do not demonstrate LTP.
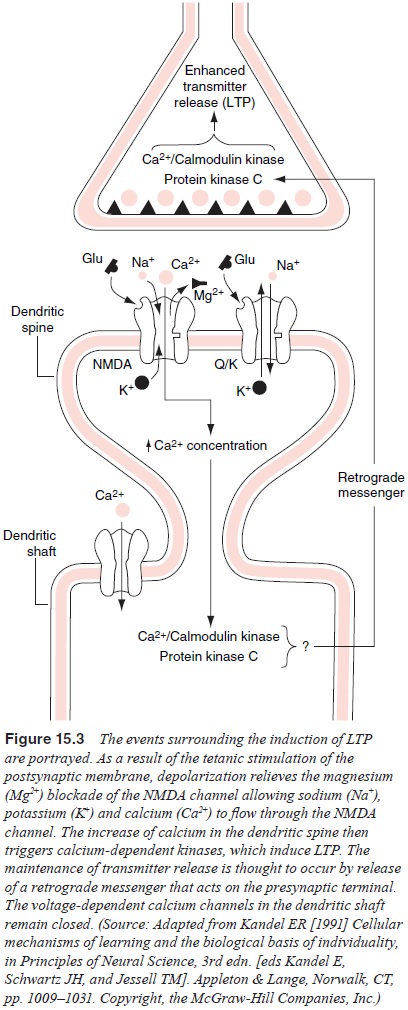
The events surrounding LTP are schematically
described here (Figure 15.3); the reader is referred to a detailed explana-tion
of the cellular and molecular aspects of LTP (Kandel, 1991Shepherd, 1994).
Studies of the CA1 region of the hippocampus reveal that LTP is
mediated by the neurotransmitter glutamate.
LTP has been stressed as a cellular mechanism of
infor-mational connectivity in the hippocampus. Brief mention should also be
made of long-term depression (LTD). LTD is the opposite of LTP and refers to
“use-dependent long-lasting decreases in synaptic strength” (Linden and Connor,
1995). LTD may have a number of advantages and work in parallel with LTP with
respect to memory functioning. LTD may help reset synapses that have been
potentiated by LTP, to prevent saturation. It may serve as a cellular mechanism
of forgetting (Tsumoto, 1993) and, finally, may also form an active inhibitory
system to attenuate signals from adjacent potentiated synapses. The specific
role of LTD in the coordination of memory is still unclear. For a detailed
de-scription of the potential cellular and molecular mechanisms in-volved in
LTD, the reader is referred to other sources (Linden and Connor, 1995; Linden,
1994). The phenomena of LTP and LTD provide an example in which neural
cytoarchitecture and the underlying cellular and molecular levels may actually
conform to the principles of association and connectivity. Computational models
have been developed to further our understanding of the manner in which
learning and memory emerge from the proper-ties of synaptic plasticity embodied
in the circuitry of the hip-pocampus (Churchland and Sejnowski, 1992; Traub and
Miles, 1991). The attraction of computational models is that they may clarify
the link between aspects of LTP and LTD within local networks of associated
neurons at the cellular level and the events at the behavioral level.
Related Topics