Chapter: Microbiology and Immunology: Morphology and Physiology of Bacteria
Cell Wall - Structure and Functions of Bacterial Cell Envelope
Structure and Functions of Bacterial Cell Envelope
The outer layer or cell envelope provides a structural and physi-ological barrier between the protoplasm (inside) of the cell and the external environment. The cell envelope protects bacteria against osmotic lysis and gives bacteria rigidity and shape. The cell envelope primarily consists of two components: a cell wall and cytoplasmic or plasma membrane. It encloses the proto-plasm, which consists of (i) cytoplasm, (ii) cytoplasmic inclu-sions (mesosomes, ribosomes, inclusion granules, vacuoles), and (iii) a single circular DNA (Fig. 2-7).
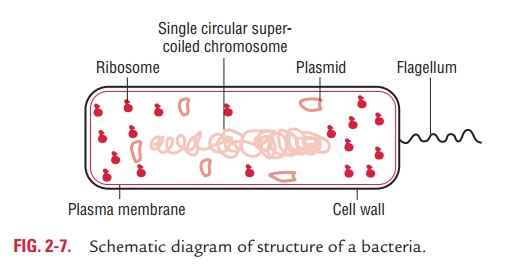
Cell Wall
Prokaryotic cells almost always are bounded by a fairly rigid and chemically complex structure present between the cell membrane and capsule/slime layer called the cell wall. Peptidoglycan is the main component of the cell wall and is responsible for the shape and strength of the cell. It is a disac-charide and contains two sugar derivatives—N-acetylglucos-amine and N-acetylmuramic acid—joined together by short peptide chains. N-acetylmuramic acid carries a tetrapeptide side chain consisting of D- and L-amino acids (D-glutamic acid and L-alanine) with mesodiaminopimelic acid (Gram-negative bacteria) or L-lysine (Gram-positive bacteria). Tetrapeptide side chains are interconnected by pentaglycine bridges. Most Gram-negative cell walls lack interpeptide bridge. Cell wall provides shape to the cell and protects bacteria from changes inosmotic pressure, which within the bacteria cell measures 5–20 atmospheres.
Bacterial cells can be classified into Gram-positive or Gram-negative based on the structural differences between Gram-positive and Gram-negative cell walls. The cell walls of the Gram-positive bacteria have simpler chemical structures compared to Gram-negative bacteria.
◗ Gram-positive cell wall
The Gram-positive cell wall is thick (15–80 nm) and more homogenous than that of the thin (2 nm) Gram-negative cell wall. The Gram-positive cell wall contains large amount of peptidoglycan present in several layers that constitutes about 40–80% of dry weight of the cell wall (Fig. 2-8). The Gram-positive
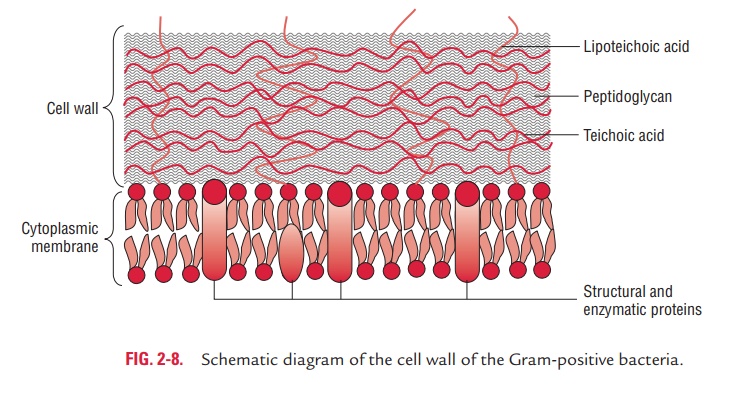
cell wall consists primarily of teichoic and teichuronic acids. These two components account for up to 50% of the dry weight of the wall and 10% of the dry weight of the total cell.
1. Teichoic acids: Teichoic acids are polymers of polyribitolphosphate or polyglycerol phosphate containing ribitol and glycerol. These polymers may have sugar or amino acid substi-tutes, either as side chain or within the chain of polymer. Teichoic acids are of two types—wall teichoic acid (WTA) and lipoteichoic acids (LTA). They are connected to the peptidoglycan by a covalent bond with the six hydroxyl of N-acetylmuramic acid in the WTA and to plasma membranelipids in LTA
2. Teichuronic acid: Teichuronic acid consists of repeat unitsof sugar acids (such as N-acetylmannuronic or D-glucuronic acid). They are synthesized in place of teichoic acids when phosphate supply to the cell is limited.
Gram-positive cell wall also contains neutral sugars (such as mannose, arabinose, rhamnose, and glucosamine) and acidic sugars (such as glucuronic acid and mannuronic acid),
which occur as subunits of polysaccharides in the cell wall.
◗ Gram-negative cell wall
The Gram-negative cell wall is much more complex than the Gram-positive cell wall. Peptidoglycan content in the Gram-negative cell wall is significantly less than the Gram-positive cell wall. Only 1–2 layers of peptidoglycan (2–8 nm) are present just outside the cell membrane. The Gram-negative cell wall outside the peptidoglycan layer contains three main components—(a) lipoprotein layer, (b) outer membrane, and (c) lipopolysaccharides (Fig. 2-9).
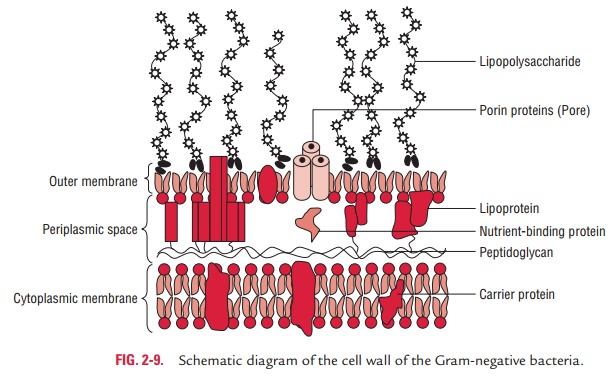
Lipoprotein layer: The lipoprotein layer is mainly composed ofBraun’s lipoprotein. Braun’s lipoprotein is a small lipoprotein that is covalently joined to the underlying peptidoglycan and embedded in the outer membrane by its hydrophobic end.
The lipoprotein stabilizes the outer membrane of the Gram-negative cell wall.
Outer membrane: The outer membrane is a bilayeredstructure; its inner part resembles in composition with that of the cell membrane, while its outer part contains a distinctive component called lipopolysaccharide. The outer membrane and plasma membrane appear to be in direct contact at many sites in the Gram-negative wall. The outer membrane has a variety of proteins as follows:
Porins: The outer membrane has special channels consist-ing of protein molecules called porins. These porins have many functions:
· They permit the passive diffusion of low-molecular weight hydrophilic compounds, such as sugars, amino acids, and certain ions;
· They exclude hydrophobic molecules; and
· They serve to protect the cell.
Outer membrane proteins (OMPs):These include thefollowing:
· Omp C, D, F, and PhoE&LamB are the four major proteins of the outer membrane that are responsible for most of the transmembrane diffusion of maltose and maltodextrins.
· Tsx, the receptor for T6 bacteriophage, is responsible for the transmembrane diffusion of nucleosides and some amino acids.
· Omp A protein anchors the outer membrane to the peptidoglycan layer. It is also the sex pilus receptor in F-mediated bacterial conjugation.
The outer membrane also contains proteins that are involved in the transport of specific molecules, such as vitamin B12 and iron-siderophore complexes; it also contains a limited num-ber of minor proteins, such as enzymes, phospholipases, and proteases.
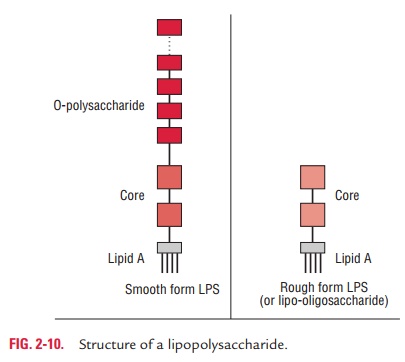
Lipopolysaccharides: Lipopolysaccharides (LPS) are complexmolecules present in the outer membrane of the Gram-negative bacteria. Structurally, the LPS consist of three main components— lipid A, the core oligosaccharide, and the O polysaccharide or O-antigen (Fig. 2-10).
· Lipid A:This consists of phosphorylated glucosamine disac-charide units, to which a number of long-chain fatty acids are attached. This also consists of hydroxymyristic acid, a unique fatty acid, which is associated with endotoxic activity of the LPS. There is a little variation in the structure of the lipid A among different species of the Gram-negative bacte-ria. However, it remains the same within the bacteria of the same species.
· Core oligosaccharide :The core oligosaccharide includes twocharacteristic sugars—ketodeoxyoctanoic acid (KDO) and a heptose—both joined together by lipid A. This is genus specific and similar in all Gram-negative bacteria. Lipoo-ligosaccharides (LOS) are smaller glycolipids. They have relatively short, multiantennary (i.e., branched) glycans present in bacteria (e.g., Neisseria meningitidis, N. gonorrhoeae, Haemophilusinfluenzae, andHaemophilusducreyi) that colo-nize mucosal surfaces. They exhibit extensive antigenic and structural diversity even within a single strain. LOS is an important virulence factor. Epitopes on LOS have a termi-nalN-acetyllactosamine (Gal(b)l-4-GlcNAc) residue, which is immunochemically similar to the precursor of the human erythrocyte i antigen. Sialylation of the N-acetyllactosamine residue in vivo provides the organism with the environmen-tal advantages of molecular mimicry of a host antigen and the biologic masking thought to be provided by sialic acids.
· polysaccharide or O-antigen:It is the portion extendingoutwards from the core. It has several peculiar sugars and varies in composition between bacterial strains, conferring species-specific antigen specificity. It is exposed to host– immune system. Gram-negative bacteria may thwart host defenses by rapidly changing the nature of their O side chains to avoid detection.
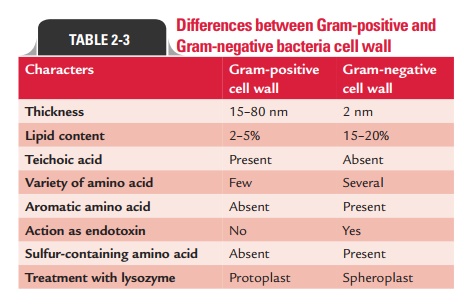
Differences between Gram-positive and Gram-negative cell walls are summarized in Table 2-3.
Periplasmic space
Periplasmic space is a distinct space between cell membrane and outer membrane (innermost layer of Gram-negative cell wall) in Gram-negative bacteria. This space is filled with a loose layer of peptidoglycan matrix. The periplasmic space of Gram-negative bacteria contains many proteins that participate in nutrient acquisition, and many hydrolytic enzymes, beta-lactamases binding proteins, and enzymes that participate in the peptidoglycan synthesis. Polymers of D-glucose, called membrane-derived oligosaccharides, appear to play a role in osmoregulation. The periplasmic space is less distinct in Gram-positive cell walls.
◗ Cell wall of acid-fast bacilli
The cell wall of acid-fast bacilli, such as M. tuberculosis, contains large amounts of waxes known as mycolic acids. The cell wall is composed of peptidoglycan and an outer asymmetric lipid bilayer. The inner lipid bilayer contains mycolic acids linked to an arabinoglycan protein and the outer layer contains other extractable lipids. This hydrophobic structure renders these bacteria resistant to many harsh chemicals including deter-gents and strong acids. During staining, if dye is introduced into these cells by brief heating or treatment with detergents, they resist decolorization by sulfuric acid or acid alcohol, and are therefore called acid-fast organisms.
◗ Atypical forms of bacteria
Atypical forms of bacteria include (i) cell wall deficient forms, (ii) pleomorphic bacteria, and (iii) involution forms. Many agents, such as antibiotics, lysozyme, and bacteriophages inter-fere or inhibit the synthesis of bacterial cell wall components, resulting in the formation of defective bacteria.
1. Cell wall deficient forms: The cell wall could be removed byhydrolysis with lysozyme or by blocking peptidoglycan synthesis with an antibiotic such as penicillin. These defective bacteria are believed to play a role in the persistence of pyelone-phritis and other chronic infections. Cell wall deficient forms without cell walls or with deficient cell walls may be of various types, such as—protoplasts, spheroplasts, mycoplasma, and L-forms.
Protoplasts:These are defective unstable forms of bacteriawith an intact cytoplasmic membrane but without any cell wall. In hypertonic media, these are produced from Gram-positive cells on treatment with lysozyme.
Spheroplasts:These are defective forms derived from Gram-negative bacteria in the presence of EDTA (ethylenediami-netetraacetic acid). The EDTA disrupts the outer membrane allowing access of lysozyme and resulting in formation of spheroplasts. Spheroplasts are osmotically fragile and still retain outer membrane and entrapped peptidoglycan.
Mycoplasma:These are naturally occurring bacteria withoutcell wall. They do not possess any definite shape. They are very minute in size measuring 50–300 nm in diameter.
L-forms:This is named after Lister Institute, London, wherethe abnormal form of Streptobacillusmoniliformis was first demonstrated. The L-forms do not exhibit any regular size and shape. They may be spherical or disc shaped, about 0.1– 20 mm in diameter. They are difficult to cultivate and usu-ally require a medium that is solidified with agar as well as having the right osmotic strength. They are produced more readily with exposure to penicillin than with lysozyme. Some bacterial species produce L-forms spontaneously. L-forms in the host may produce chronic infections and are relatively resistant to antibiotic treatment
2. Pleomorphic bacteria: Pleomorphic bacteria (e.g.,Yersiniapestis) may show considerable variation in size and shape calledpleomorphism.
3. Involution forms: The involution forms are those that onageing of culture show swollen and aberrant forms, especially in high salt concentration.
◗ Demonstration of cell wall
The cell walls can be demonstrated by (a) differential staining procedure, (b) electron microscopy, (c) plasmolysis, (d) micro-dissection, (e) mechanical rupture of the cell, and (f) serological test by exposure to specific antibodies.
Related Topics