Chapter: Medical Physiology: Digestion and Absorption in the Gastrointestinal Tract
Absorption in the Small Intestine
Absorption in the Small Intestine
Absorption from the small intestine each day consists of several hundred grams of carbohydrates, 100 or
The absorptive capacity of the normal small intestine is far greater than this: as much as several kilograms of car-bohydrates per day, 500 grams of fat per day, 500 to 700 grams of proteins per day, and 20 or more liters of water per day. The large intestine can absorb still addi-tional water and ions, although very few nutrients.
Absorption of Water
Isosmotic Absorption. Water is transported through theintestinal membrane entirely by diffusion. Further-more, this diffusion obeys the usual laws of osmosis. Therefore, when the chyme is dilute enough, water is absorbed through the intestinal mucosa into the blood of the villi almost entirely by osmosis.
Conversely, water can also be transported in the opposite direction—from plasma into the chyme. This occurs especially when hyperosmotic solutions are dis-charged from the stomach into the duodenum. Within minutes, sufficient water usually will be transferred by osmosis to make the chyme isosmotic with the plasma.
Absorption of Ions
Active Transport of Sodium. Twenty to 30 grams of sodiumare secreted in the intestinal secretions each day. In addition, the average person eats 5 to 8 grams of sodium each day. Therefore, to prevent net loss of sodium into the feces, the intestines must absorb 25 to 35 grams of sodium each day, which is equal to about one seventh of all the sodium present in the body.
Whenever significant amounts of intestinal secre-tions are lost to the exterior, as in extreme diarrhea, the sodium reserves of the body can sometimes be depleted to lethal levels within hours. Normally, however, less than 0.5 per cent of the intestinal sodium is lost in the feces each day because it is rapidly absorbed through the intestinal mucosa. Sodium also plays an important role in helping to absorb sugars and amino acids, as we shall see in subsequent discussions.
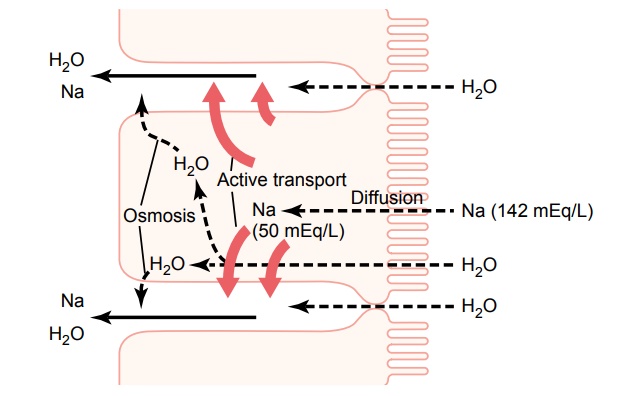
The basic mechanism of sodium absorption from the intestine is shown in Figure 65–8. The principles of this mechanism are also essentially the same as for absorption of sodium from the gallbladder and renal tubules.
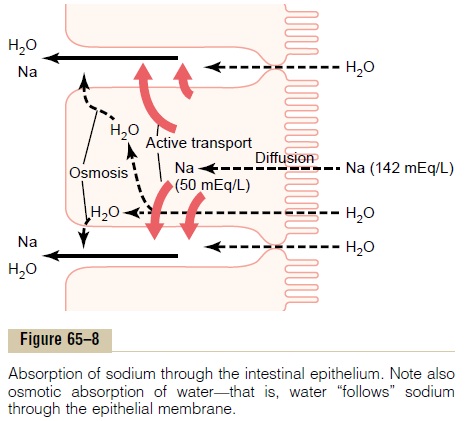
The motive power for sodium absorption is pro-vided by active transport of sodium from inside the epithelial cells through the basal and side walls of these cells into paracellular spaces. This is demon-strated by the heavy red arrows in Figure 65–8. This active transport obeys the usual laws of active trans-port: it requires energy, and the energy process is cat-alyzed by appropriate adenosine triphosphatase enzymes in the cell membrane Part of the sodium is absorbed along with chloride ions; in fact, the negatively charged chloride ions are mainly passively “dragged” by the positive electrical charges of the sodium ions.
Active transport of sodium through the basolateral membranes of the cell reduces the sodium concentra-tion inside the cell to a low value (about 50 mEq/L), as also shown in Figure 65–8. Because the sodium con-centration in the chyme is normally about 142 mEq/L (that is, about equal to that in plasma), sodium moves down this steep electrochemical gradient from the chyme through the brush border of the epithelial cell into the epithelial cell cytoplasm. This provides still more sodium ions to be transported by the epithelial cells into the paracellular spaces.
Osmosis of the Water. The next step in the transportprocess is osmosis of water into the paracellular spaces. This occurs because a large osmotic gradient has been created by the elevated concentration of ions in the paracellular space. Much of this osmosis occurs through the tight junctions between the apical borders of the epithelial cells, but much also occurs through the cells themselves. And osmotic movement of water creates flow of fluid into and through the paracellular spaces and, finally, into the circulating blood of the villus.
Aldosterone Greatly Enhances Sodium Absorption. When aperson becomes dehydrated, large amounts of aldos-terone almost always are secreted by the cortices of the adrenal glands. Within 1 to 3 hours this aldosterone causes increased activation of the enzyme and trans-port mechanisms for all aspects of sodium absorption by the intestinal epithelium. And the increased sodium absorption in turn causes secondary increases in absorption of chloride ions, water, and some other substances.
This effect of aldosterone is especially important in the colon because it allows virtually no loss of sodium chloride in the feces and also little water loss. Thus, the function of aldosterone in the intestinal tract is the same as that achieved by aldosterone in the renal tubules, which also serves to conserve sodium chloride and water in the body when a person becomes dehydrated.
Absorption of Chloride Ions in the Duodenum and Jejunum. In the upper part of the small intestine, chloride ion absorption is rapid and occurs mainly by diffusion— that is, absorption of sodium ions through the epithe-lium creates electronegativity in the chyme and electropositivity in the paracellular spaces between the epithelial cells. Then chloride ions move along this electrical gradient to “follow” the sodium ions.
Absorption of Bicarbonate Ions in the Duodenum and Jejunum.
Often large quantities of bicarbonate ions must be reabsorbed from the upper small intestine because large amounts of bicarbonate ions have been secreted into the duodenum in both pancreatic secretion and bile. The bicarbonate ion is absorbed in an indirect way as follows: When sodium ions are absorbed, moderate amounts of hydrogen ions are secreted into the lumen of the gut in exchange for some of the sodium. These hydrogen ions in turn combine with the bicarbonate ions to form carbonic acid (H2CO3), which then disso-ciates to form water and carbon dioxide. The water remains as part of the chyme in the intestines, but the carbon dioxide is readily absorbed into the blood and subsequently expired through the lungs. Thus, this is so-called “active absorption of bicarbonate ions.” It is the same mechanism that occurs in the tubules of the kidneys.
Secretion of Bicarbonate Ions in the Ileum and Large Intestine—Simultaneous Absorption of Chloride Ions
The epithelial cells on the surfaces of the villi in the ileum as well as on all surfaces of the large intestine have a special capability of secreting bicarbonate ions in exchange for absorption of chloride ions. This is important because it provides alkaline bicarbonate ions that neutralize acid products formed by bacteria in the large intestine.
Extreme Secretion of Chloride Ions, Sodium Ions, and Water fromthe Large Intestine Epithelium in Some Types of Diarrhea. Deepin the spaces between the intestinal epithelial folds are immature epithelial cells that continually divide to form new epithelial cells. These in turn spread outward over the luminal surfaces of the intestines. While still in the deep folds, the epithelial cells secrete sodium chloride and water into the intestinal lumen. This secretion in turn is reabsorbed by the older epithelial cells outside the folds, thus providing flow of water for absorbing intestinal digestates.
The toxins of cholera and of some other types of diar-rheal bacteria can stimulate the fold secretion so greatly that this secretion often becomes much greater than can be reabsorbed, thus sometimes causing loss of 5 to 10 liters of water and sodium chloride as diarrhea each day. Within 1 to 5 days, many severely affected patients die from this loss of fluid alone.
Extreme diarrheal secretion is initiated by entry of a subunit of cholera toxin into the epithelial cells. This stimulates formation of excess cyclic adenosine monophosphate, which opens tremendous numbers of chloride channels, allowing chloride ions to flow rapidly from inside the cell into the intestinal crypts. In turn, this is believed to activate a sodium pump that pumps sodium ions into the crypts to go along with the chlo-ride ions. Finally, all this extra sodium chloride causes extreme osmosis of water from the blood, thus provid-ing rapid flow of fluid along with the salt. All this excess fluid washes away most of the bacteria and is of value in combating the disease, but too much of a good thing can be lethal because of serious dehydration of the whole body that might ensue. In most instances, the life of a cholera victim can be saved by administration of tremendous amounts of sodium chloride solution to make up for the loss.
Absorption of Other Ions. Calcium ionsare activelyabsorbed into the blood especially from the duode-num, and the amount of calcium ion absorption is very exactly controlled to supply exactly the daily need of the body for calcium. One important factor controlling calcium absorption is parathyroid hormone secreted by the parathyroid glands, and another is vitamin D. Parathyroid hormone activates vitamin D, and the activated vitamin D in turn greatly enhances calcium absorption.
Iron ions are also actively absorbed from the smallintestine. The principles of iron absorption and regu-lation of its absorption in proportion to the body’s need for iron, especially for the formation of hemo-globin.
Potassium, magnesium, phosphate, and probably still other ions can also be actively absorbed through theintestinal mucosa. In general, the monovalent ions are absorbed with ease and in great quantities. Conversely, bivalent ions are normally absorbed in only small amounts; for example, maximum absorption of calcium ions is only 1/50 as great as the normal absorp-tion of sodium ions. Fortunately, only small quantities of the bivalent ions are normally required daily by the body.
Absorption of Nutrients
Absorption of Carbohydrates
Essentially all the carbohydrates in the food are absorbed in the form of monosaccharides; only a small fraction are absorbed as disaccharides and almost none as larger carbohydrate compounds. By far the most abundant of the absorbed monosaccharides is glucose, usually accounting for more than 80 per cent of carbohydrate calories absorbed. The reason for this is that glucose is the final digestion product of our most abundant carbohydrate food, the starches. The remaining 20 per cent of absorbed monosaccharides are composed almost entirely of galactose and fruc-tose, the galactose derived from milk and the fructoseas one of the monosaccharides digested from cane sugar.
Virtually all the monosaccharides are absorbed by an active transport process. Let us first discuss the absorption of glucose.
Glucose Is Transported by a Sodium Co-Transport Mechanism.
In the absence of sodium transport through the intes-tinal membrane, virtually no glucose can be absorbed. The reason is that glucose absorption occurs in a co-transport mode with active transport of sodium.
There are two stages in the transport of sodium through the intestinal membrane. First is active trans-port of sodium ions through the basolateral mem-branes of the intestinal epithelial cells into the blood, thereby depleting sodium inside the epithelial cells. Second, decrease of sodium inside the cells causes sodium from the intestinal lumen to move through the brush border of the epithelial cells to the cell interiors by a process of facilitated diffusion. That is, a sodium ion combines with a transport protein, but the trans-port protein will not transport the sodium to the inte-rior of the cell until the protein itself also combines with some other appropriate substance such as glucose. Fortunately, intestinal glucose also combines simultaneously with the same transport protein, and then both the sodium ion and glucose molecule are transported together to the interior of the cell. Thus, the low concentration of sodium inside the cell liter-ally “drags” sodium to the interior of the cell and along with it the glucose at the same time. Once inside the epithelial cell, other transport proteins and enzymes cause facilitated diffusion of the glucose through the cell’s basolateral membrane into the paracellular space and from there into the blood.
To summarize, it is the initial active transport of sodium through the basolateral membranes of the intestinal epithelial cells that provides the eventual motive force for moving glucose also through the membranes.
Absorption of Other Monosaccharides. Galactose is trans-ported by almost exactly the same mechanism as glucose. Conversely, fructose transport does not occur by the sodium co-transport mechanism. Instead, fruc-tose is transported by facilitated diffusion all the way through the intestinal epithelium but not coupled with sodium transport.
Much of the fructose, on entering the cell, becomes phosphorylated, then converted to glucose, and finally transported in the form of glucose the rest of the way into the blood. Because fructose is not co-transported with sodium, its overall rate of transport is only about one half that of glucose or galactose.
Absorption of Proteins
Most proteins, after digestion, are absorbed through the luminal membranes of the intestinal epithelial cells in the form of dipeptides, tripeptides, and a few free amino acids. The energy for most of this transport is supplied by a sodium co-transport mechanism in the same way that sodium co-transport of glucose occurs. That is, most peptide or amino acid molecules bind in the cell’s microvillus membrane with a specific transport protein that requires sodium binding before transport can occur. After binding, the sodium ion then moves down its electrochemical gradient to the interior of the cell and pulls the amino acid or peptide along with it. This is called co-transport (or secondary active transport) of the amino acids and peptides. A few amino acids donot require this sodium co-transport mechanism but instead are transported by special membrane trans-port proteins in the same way that fructose is trans-ported, by facilitated diffusion.
At least five types of transport proteins for trans-porting amino acids and peptides have been found in the luminal membranes of intestinal epithelial cells. This multiplicity of transport proteins is required because of the diverse binding properties of different amino acids and peptides.
Absorption of Fats
Earlier, it was pointed out that when fats are digested to form monoglycerides and free fatty acids, both of these digestive end products first become dissolved in the central lipid portions of bile micelles. Because the molecular dimensions of these micelles are only 3 to 6 nanometers in diameter, and because of their highly charged exterior, they are soluble in chyme. In this form, the monoglycerides and free fatty acids are carried to the surfaces of the microvilli of the intestinal cell brush border and then penetrate into the recesses among the moving, agitating microvilli. Here, both the monoglycerides and fatty acids diffuse imme-diately out of the micelles and into the interior of the epithelial cells, which is possible because the lipids are also soluble in the epithelial cell membrane. This leaves the bile micelles still in the chyme, where they function again and again to help absorb still more monoglycerides and fatty acids.
Thus, the micelles perform a “ferrying” function that is highly important for fat absorption. In the presence of an abundance of bile micelles, about 97 per cent of the fat is absorbed; in the absence of the bile micelles, only 40 to 50 per cent can be absorbed.
After entering the epithelial cell, the fatty acids and monoglycerides are taken up by the cell’s smooth endoplasmic reticulum; here, they are mainly used to form new triglycerides that are subsequently released in the form of chylomicrons through the base of the epithelial cell, to flow upward through the thoracic lymph duct and empty into the circulating blood.
Direct Absorption of Fatty Acids into the Portal Blood. Smallquantities of short- and medium-chain fatty acids, such as those from butterfat, are absorbed directly into the portal blood rather than being converted into triglyc-erides and absorbed by way of the lymphatics. The cause of this difference between short- and long-chain fatty acid absorption is that the short-chain fatty acids are more water-soluble and mostly are not recon-verted into triglycerides by the endoplasmic reticulum. This allows direct diffusion of these short-chain fatty acids from the intestinal epithelial cells directly into the capillary blood of the intestinal villi.
Related Topics