Chapter: Medical Microbiology: An Introduction to Infectious Diseases: Mycobacteria
Mycobacterium - Bacteriology
MYCOBACTERIUM
BACTERIOLOGY
MORPHOLOGY AND STRUCTURE
The mycobacteria are slim, Gram-positive bacilli (0.2 – 0.4 x 2 – 10 μm). They are nonmotile, obligate aerobes that do not form spores. The cell wall contains peptidoglycan similar to that of other Gram-positive organisms, except that it contains Nglycolylmuramic, rather than N-acetylmuramic, acid. Attached to peptidoglycan are a myriad of branched chain polysaccharides, proteins, and lipids. Of particular importance are long-chain fatty acids called mycolic acids. The mycolic acids, for which themycobacteria are named, make up more than 60% of the total cell wall mass and are distinctive for each species. Other lipid components include mycosides, sulfolipids, and lipoarabinomannan (LAM), a complex molecule extending from the plasma membrane to the surface. LAM is structurally and functionally analogous to the lipopolysaccharide of Gram-negative bacteria. Porin and other proteins are found throughout the cell wall.
The cell wall lipids make the cell surface hydrophobic, rendering mycobacteria resis-tant to staining with basic aniline dyes unless they are applied with heat or detergents, or for prolonged periods of time. Once stained, however, mycobacteria resist decolorization with a mixture of 3% hydrochloric acid and 95% ethanol. These properties are described as acid fastness or, more properly, acid – alcohol fastness, and the bacteria possessing them are called acid-fast bacilli. This characteristic allows mycobacteria to be readily distinguished from other genera by microscopic exami-nation of smears stained with carbol fuchsin (Ziehl – Neelsen/Kinyoun techniques), or with the more recently introduced fluorochromes (auramine – rhodamine). Organisms stained with the latter reagents fluoresce brightly when viewed through an appropriate microscope, making the organisms more visually apparent and, thus, decreasing the time required for their detection.
GROWTH
The most important pathogen, M. tuberculosis, shows enhanced growth in 10% carbon dioxide and at a pH of about 6.5 to 6.8. Nutritional requirements vary among species and range from the ability of some nonpathogens to multiply on the washers of water faucets to the strict intracellular parasitism of M. leprae, which does not grow in artifi-cial media or cell culture. Mycobacteria grow more slowly than most human patho-genic bacteria because of their hydrophobic cell surface, which causes them to clump and inhibits permeability of nutrients into the cell. Addition of a surfactant (Tween 80) to cultures of M. tuberculosis wets the surface and leads to dispersed and more rapid growth.
CLASSIFICATION
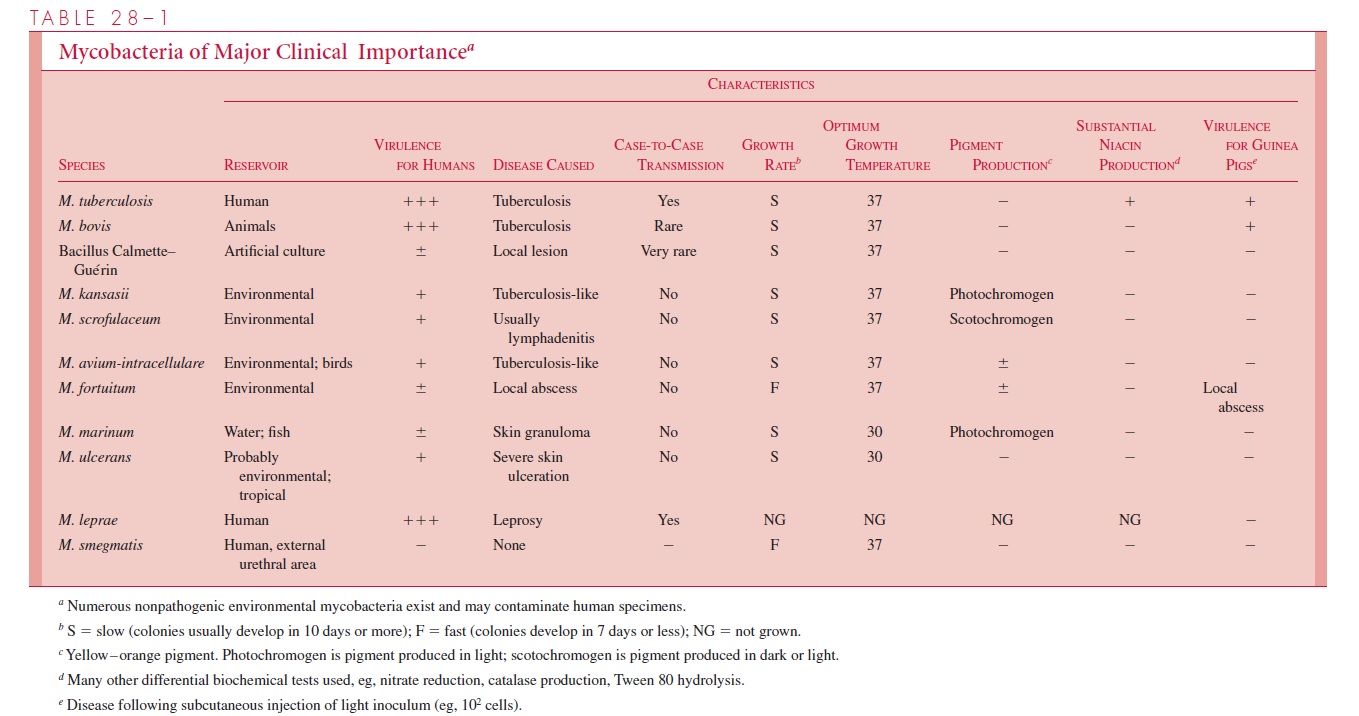
Until recently, mycobacterial classification has been based on a constellation of phe-notypic characteristics, including nutritional and temperature requirements, growth rates, pigmentation of colonies grown in light or darkness, key biochemical tests, the cellular constellation of free fatty acids, and the range of pathogenicity in experimental animals. Some of the more important characteristics are summarized in Table 28 – 1. Increasingly, this classification system is yielding to molecular-based techniques. The identification of species-specific rRNA and DNA sequences has resulted in the revision and expansion of the older phenotype-based classification system, and the provision of an increasing array of species-specific DNA probes to clinical mycobacteriology laboratories.
Related Topics