Chapter: Human Neuroanatomy(Fundamental and Clinical): Internal Capsule and Commissures of the Brain
The Visual Pathway of the Brain
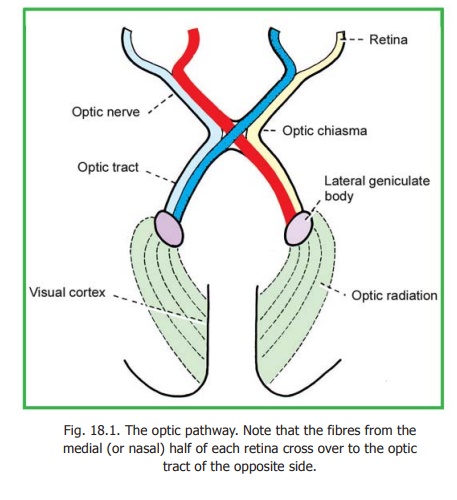
The Visual Pathway
The peripheral receptors for light are situated in the retina. Nerve fibres arising in the retina constitute the optic nerve. The right and left optic nerves join to form the optic chiasma in which many of their fibres cross to the opposite side. The uncrossed fibres of the optic nerve, along with the fibres that have crossed over from the opposite side form theoptic tract.
The optic tract terminates predominantly in the lateral geniculate body. Fresh fibres arising in this body form the geniculo-calcarine tract or optic radiation which ends in the visual areas of the cerebral cortex (Fig. 18.1).
The Retina
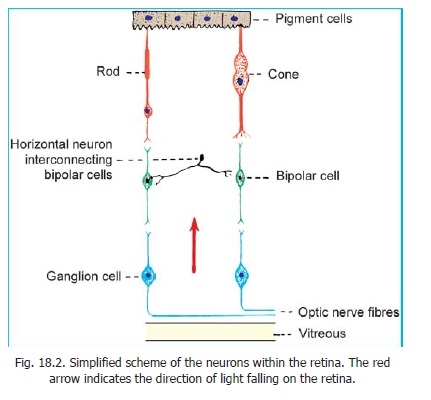
The retina has a complex structure. It contains photoreceptors that convert the stimulus of light into nervous impulses. These receptors are of two kinds, rods and cones (Fig. 18.2). There are about seven million cones in each retina. The rods are far more numerous, and number more than 100 million.
The cones respond best to bright light. They are responsible for sharp vision and for discrimination of colour. They are most numerous in the central region of the retina which is responsible for sharp vision. This area is about 6mm in diameter. Within this region there is a yellowish area called the macula lutea. The centre of the macula shows a small depression called the fovea centralis.
Medial to the central area, there is a circular area called the optic disc. This area is devoid of photoreceptors and is, therefore, called the blind spot. The fibres of the optic nerve leave the eyeball through the region of the optic disc.
The macula lies exactly in the optical axis of the eyeball. When any object is viewed critically its image is formed on the macula. The fovea centralis is believed to contain cones only. Rods, on the other hand, predominate in peripheral parts of the retina. They can respond to poor light and specially to movement across the field of vision.
Each rod or cone may be regarded as a modified neuron. It consists of a cell body, a peripheral process and a central process. The cell body contains a nucleus. The peripheral process is rod shaped in the case of rods, and cone shaped in the case of cones (hence the names rods and cones). The ends of these peripheral processes are separated from one another by processes of pigment cells. The central processes of rods and cones are like those of neurons. They end by synapsing with other neurons within the retina. For details of the structure of rods and cones, see the author’s HUMAN HISTOLOGY.
The basic neuronal arrangement within the retina is shown in Fig. 18.2. The central processes of rods and cones synapse with the peripheral processes of bipolar cells. The central processes of bipolar cells synapse with dendrites of ganglion cells. Axons arising from ganglion cells form the fibres of the optic nerve.
The various elements mentioned above form a series of layers within the retina. The outermost layer (towards the choroid) is formed by the pigments cells, followed in sequence by the rods and cones, the bipolar cells, the ganglion cells, and a layer of optic nerve fibres. The layer of optic nerve fibres is apposed to the vitreous. It will be obvious that light has to pass through several of the layers of the retina to reach the rods and cones. This ‘inverted’ arrangement of the retina is necessary as passage of light in the reverse direction would be obstructed by the layer of pigment cells.
The pigment cells are important in spacing the rods and cones and in providing them with mechanical support. They absorb light and prevent back reflection. A nutritive and phagocytic role has also been attributed to them.
Advanced:
At this stage it must be emphasised that the structure of the retina is very much more complex than the highly simplified description above would suggest. Some points worthy of note are as follows.
1. Rods and cones are of various types. Intermediate forms between them are also present. The rods and cones have a complicated ultrastructure. Their outer segments contain photopigments which appear to be concerned in converting the stimulus of light into a nervous impulse.
2. Bipolar cells and ganglion cells are of various types. They vary in size and in their connections. Some bipolar cells are connected with cones, some with rods, and others with both rods and cones.
3. The vertically orientated elements of the retina (i.e., rods and cones, bipolar cells, ganglion cells) are interconnected by horizontally disposed neurons. Some of these appear to be devoid of axons and are called amacrine cells.
4. The number of rods and cones (over 100 million) greatly exceeds the number of ganglion cells (about one million). It is, therefore, obvious that the impulses from several rods and cones must be concentrated on a single ganglion cell; and must, therefore, travel along a single optic nerve fibre. The one to one relationship between rods and cones, bipolar cells and ganglion cells shown in Fig. 18.2 is only for sake of simplicity.
5. According to some authorities, the retina receives some fibres through the optic nerve although their source is uncertain. Other authorities however, deny the presence of such fibres.
6. Besides the nervous elements, the retina contains supporting cells which are believed to represent a specialised type of neuroglia.
The Visual Field and Retinal Quadrants
When the head and eyes are maintained in a fixed position, and one eye is closed, the area seen by that eye constitutes the visual field for that eye. Now if the other eye is also opened the area seen is more or less the same as was seen with one eye. In other words the visual fields of the two eyes overlap to a very great extent. On either side, however, there is a small area seen only by the eye of that side. Although the two eyes view the same area, the relative position of objects within the area appears somewhat dissimilar to the two eyes as they view the object from slightly different angles. The difference though slight, is of considerable importance as it forms the basis for the perception of depth (stereoscopic vision).
For convenience of description, the visual field is divided into right and left halves. It may also be divided into upper and lower halves so that the visual field can be said to consist of four quadrants (Fig. 18.3). In a similar manner each retina can also be divided into quadrants. Images of objects in
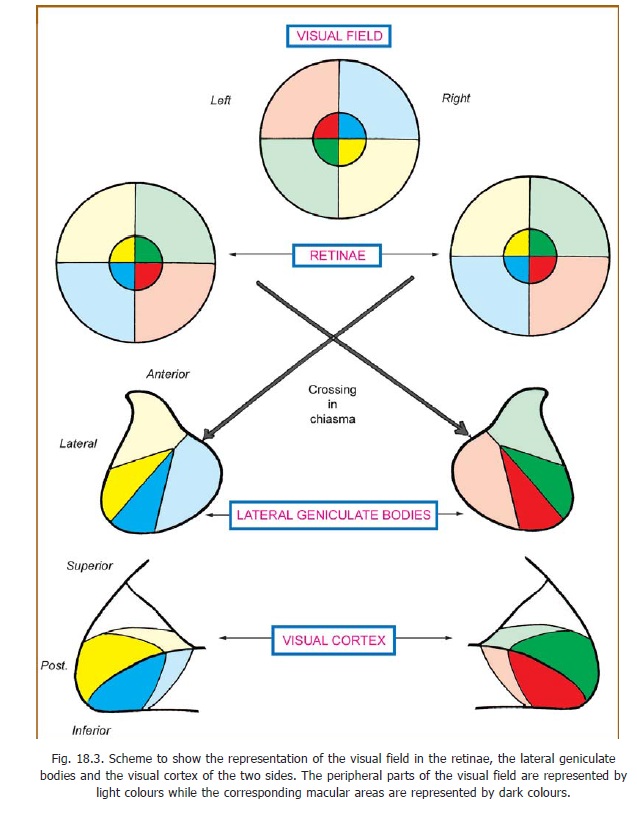
the field of vision are formed on the retina by the lens of the eyeball. As with any convex lens the image is inverted. If an object is placed in the right half of the field of vision its image is formed on the left half of the retina and vice versa. Unfortunately, instead of using the words right and left, the two halves of the retina are usually referred to as nasal (= medial) and temporal (= lateral) halves. This introduces a complication as the left half of the left eye is the temporal half, while in the case of the right eye it is the nasal half. Thus the image of an object placed in the right half of the field of vision falls on the temporal half of the left retina, and on the nasal half of the right retina.
Optic Nerve, Optic Chiasma & Optic Tract
The optic nerve is made up of axons of the ganglion cells of the retina. These axons are at first unmyelinated. The fibres from all parts of the retina converge on the optic disc. In this region the sclera has numerous small apertures and is, therefore, called the lamina cribrosa (crib = sieve). Bundles of optic nerve fibres pass through these apertures. Each fibre acquires a myelin sheath as soon as it pierces the sclera. The fibres of the nerve arising from the four quadrants of the retina maintain the same relative position within the nerve. The fibres arising from the macula are numerous and form the papillomacular bundle. Close to the eyeball, the macular fibres occupy the lateral part of the nerve, but by the time they reach the chiasma they lie in the central part of the nerve.
The fibres of the optic nerve arising in the nasal half of each retina enter the optic tract of the opposite side after crossing in the chiasma. Fibres from the temporal half of each retina enter the optic tract of the same side (Fig. 18.1). Thus the right optic tract comes to contain fibres from the right halves of both retinae, and the left tract from the left halves. In other words, all optic nerve fibres carrying impulses relating to the left half of the field of vision are brought together in the right optic tract and vice versa. We have already noted that each optic tract carries these fibres to the lateral geniculate body of the corresponding side.
The Lateral Geniculate Body
Some aspects of the structure of the lateral geniculate body have been considered. It has been seen that the grey matter of this body is split into six laminae (Fig. 13.17). Fibres from the eye of the same side end in laminae 2, 3, and 5; while those from the opposite eye end in laminae 1, 4, and 6. The macular fibres end in the central and posterior part of the lateral geniculate body, and this area is relatively large (Fig. 18.3). Fibres from the peripheral parts of the retina end in the anterior part of the lateral geniculate body. The upper half of the retina is represented laterally, and the lower half of the retina is represented medially. Specific points on the retina project to specific points in the lateral geniculate body. In turn, specific points of this body project to specific points in the visual cortex. In this way a point to point relationship is maintained between the retinae and the visual cortex.
Recent studies of synaptic patterns within the lateral geniculate body indicate that this nucleus is not to be regarded as a simple relay station and that various influences may modify the conduction of impulses through it. In this connection it may be noted that the lateral geniculate body receives afferents from the visual cortex.
Geniculocalcarine Tract and Visual Cortex
Fibres arising from cells of the lateral geniculate body constitute the geniculocalcarine tract or optic radiation. These fibres pass through the retrolentiform part of the internal capsule. The optic radiation ends in the visual areas of the cerebral cortex (areas 17, 18, 19).
Some fibres of the radiation loop forwards and downwards into the temporal lobe before turning backwards into the occipital lobe. The highest fibres may loop into the parietal lobe.
The cortex of each hemisphere receives impulses from the retinal halves of the same side (i.e., from the opposite half of the field of vision). The upper quadrants of the retina are represented above the calcarine sulcus, and the lower quadrants below it. The cortical area for the macula is larger than that for peripheral areas. It occupies the posterior part of the visual area. The cortical area for the peripheral part of the retina is situated anterior to the area for the macula.
Clinical:
Injuries to different parts of the visual pathway can produce various kinds of defects. Loss of vision in one half (right or left) of the visual field is called hemianopia. If the same half of the visual field is lost in both eyes the defect is said to be homonymous and if different halves are lost the defect is said to be heteronymous. Note that the hemianopia is named in relation to the visual field and not to the retina.
Injury to the optic nerve will obviously produce total blindness in the eye concerned. Damage to the central part of the optic chiasma (e.g., by pressure from an enlarged hypophysis) interrupts the crossing fibres derived from the nasal halves of the two retinae resulting in bitemporalheteronymous hemianopia. It has been claimed that macular fibres are more susceptible todamage by pressure than peripheral fibres and are affected first. When the lateral part of the chiasma is affected a nasal hemianopia results. This may be unilateral or bilateral. Complete destruction of the optic tract, the lateral geniculate body, the optic radiation or the visual cortex of one side, results in loss of the opposite half of the field of vision. A lesion on the right side leads to left homonymoushemianopia.
Partial injury may affect only one quadrant. The resulting condition is calledquadrantic anopia. Injury to looping fibres of the geniculocalcarine tract may lead to blindness in the upper quadrant when the lesion is in the temporal lobe; and in the lower quadrant when the lesion is in the parietal lobe.
It may be noted that in lesions of the visual pathway macular vision is often spared. This is so because of the large size of the macular area, and because some areas have a double blood supply (from posterior and middle cerebral arteries).
Lesions confined to areas 18 and 19 (peristriate and parastriate areas) may not result in loss of vision, but the patient may be unable to interpret images seen. This is visual agnosia.
Lesions anterior to the lateral geniculate body also interrupt fibres responsible for thepupillarylight reflex .
Related Topics