Chapter: 11th Botany : Chapter 6 : Cell: The Unit of Life
Structure and Types of chromosome

Chromosomes
Strasburger
(1875)
first reported its present in
eukaryotic cell and the term ‘chromosome’ was introduced by Waldeyer in 1888. Bridges (1916) first proved that chromosomes are the physical
carriers of genes. It is made up of DNA and associated proteins.
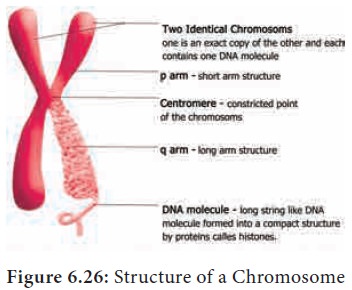
Structure of chromosome
The chromosomes are composed of thread like strands
called chromatin which is made up of
DNA, protein and RNA. Each chromosome consists of two symmetrical structures
called chromatids. During cell
division the chromatids forms well organized chromosomes with definite size and
shape. They are identical and are called sister
chromatids. A typical chromosome has narrow zones called constrictions. There are two types of
constrictions namely primary constriction and secondary constriction. The primary constriction is made up of centromere and kinetochore. Both the chromatids are united at centromere,
whose number varies. The monocentric
chromosome has one centromere and the polycentric
chromosome has many centromeres. The centromere contains a complex system of
protein fibres called kinetochore.
Kinetochore is the region of
chromosome which is attached to the spindle fibre during mitosis.
Besides primary there are secondary constrictions, represented
with few occurrence. Nucleoli
develop from these secondary constrictions are called nucleolar organizers.
Secondary constrictions contains the
genes for ribosomal RNA which induce the formation of nucleoli and are called nucleolar organizer regions (Figure
6.26).
A satellite
or SAT Chromosome are short chromosomal segment or rounded body separated from
main chromosome by a relatively elongated secondary constriction. It is a
morphological entity in certain chromosomes.
Chromonema fiber: It is a chromatin
fibre, 100 – 130 nm in diameter thought to be an element of higher order
packing of chromatin within chromosome. During prophase the chromosomal
material becomes visible as very thin filaments called chromonemata, which is
called as chromatids in early stages of condensation. Chromatid and chromonema
are the two names for the same structure a single linear DNA molecule with its
associated proteins.
Chromomeres: Chromomeres are bead like accumulations of chromatin material which are visible
along interphase chromosomes. They can be seen in polytene chromosomes. At
metaphase they are not visible.
Based on the position of centromere, chromosomes
are called telocentric (terminal
centromere), Acrocentric (terminal
centromere capped by telomere), Sub
metacentric (centromere subterminal)
and Metacentric (centromere
median). The eukaryotic chromosomes may be rod shaped (telocentric and
acrocentric), L-shaped (sub-metacentric) and V-shaped (metacentric) (Figure
6.27).

Telomere is the terminal part of chromosome. It offers stability to the chromosome. DNA of the telomere has specific sequence of nucleotides. Telomere in all eukaryotes are composed of many repeats of short DNA sequences (5’TTAGGG3’ sequence in Neurospora crassa and human beings). Maintenance of telomeres appears to be an important factor in determining the life span and reproductive capacity of cells so studies of telomeres and telomerase have the promise of providing new insights into conditions such as ageing and cancer. Telomeres prevents the fusion of chromosomal ends with one another.
Holocentric chromosomes have
centromere activity distributed along the whole surface of the chromosome
during mitosis. Holocentric condition can be seen in Caenorhabditis elegans (nematode) and many insects. There are three
types of centromere in eukaryotes. They are as follows:
Point
centromere: the type of centromere
in which the kinetochore is assembled as a result of protein recognition of
specific DNA sequences. Kinetochores assembled on point centromere bind a
single microtubule. It is also called as localized
centromere. It occurs in budding yeasts
Regional
centromere: In regional centromere
where the kinetochore is assembled on a variable array of repeated DNA
sequences. Kinetochore assembled on regional centromeres bind multiple
microtubules. It occurs in fission yeast cell, humans and so on.
Holocentromere- The
microtubules bind all along the
mitotic chromosome. Example: Caenorhabditis
elegans (nematode) and many insects.
Based on the functions of chromosome it can be
divided into autosomes and sex chromosomes.
Autosomes are present in all cells controlling
somatic characteristics of an organism. In human diploid cell, 44 chromosomes
are autosomes whereas two are sex chromosomes. Sex chromosomes are involved in
the determination of sex.
Special types of chromosomes are found only in
certain special tissues. These chromosomes are larger in size and are called giant chromosomes in certain plants and
they are found in the suspensors of the embryo. The polytene chromosome and
lamp brush chromosome occur in animals and are also called as giant chromosomes.
Polytene
chromosomes observed in the
salivary glands of Drosophila (fruit fly) by C.G. Balbiani in 1881. In larvae of many flies, midges (Dipthera)
and some insects the interphase chromosomes duplicates and reduplicates without
nuclear division. A single chromosome which is present in multiple copies form
a structure called polytene chromosome which
can be seen in light microscope.
They are genetically active. There is a distinct alternating dark bands and
light inter-bands. About 95% of DNA are present in bands and 5% in inter-bands.
The polytene chromosome has extremely large puff called Balbiani rings which is seen in Chironomous larvae. It is also
known as chromosomal puff. Puffing
of bands are the sites of intense RNA synthesis. As this chromosome occurs in
the salivary gland it is known as salivary
gland chromosomes. Polyteny is
achieved by repeated replication of
chromosomal DNA several times without nuclear division and the daughter
chromatids aligned side by side and do not separate (called endomitosis). Gene expression,
transcription of genes and RNA synthesis occurs in the bands along the polytene
chromosomes. Maternal and paternal homologues remain associated side by side is
called somatic pairing.
Lampbrush
chromosomes occur at the
diplotene stage of first meiotic prophase in oocytes of an animal Salamandar and in giant nucleus of the
unicellular alga Acetabularia. It was
first observed by Flemming in 1882. The highly condensed chromosome forms the chromosomal
axis, from which lateral loops of DNA extend as a result of intense RNA
synthesis.
Related Topics