Chapter: Medical Physiology: Pituitary Hormones and Their Control by the Hypothalamus
Posterior Pituitary Gland and Its Relation to the Hypothalamus
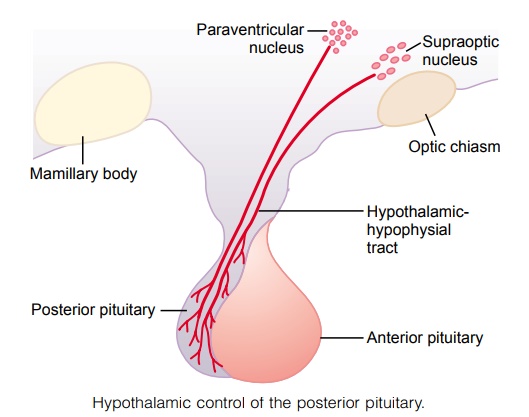
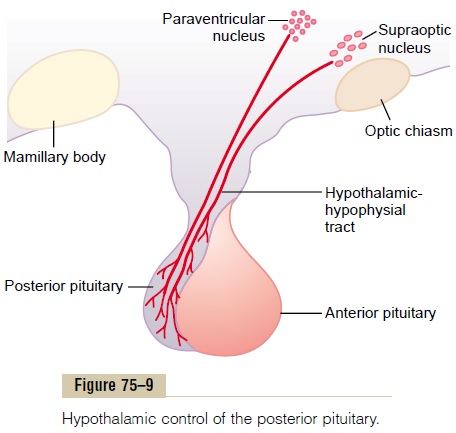
Posterior Pituitary Gland and Its Relation to the Hypothalamus
The posterior pituitary gland, also called the neurohy-pophysis, is composed mainly of glial-like cells called pituicytes. The pituicytes do not secrete hormones; theyact simply as a supporting structure for large numbers of terminal nerve fibers and terminal nerve endings from nerve tracts that originate in the supraoptic and paraventricular nuclei of the hypothalamus, as shownin Figure 75–9. These tracts pass to the neurohypoph-ysis through the pituitary stalk (hypophysial stalk). The nerve endings are bulbous knobs that contain many secretory granules. These endings lie on the surfaces of capillaries, where they secrete two posterior pituitary hormones: (1) antidiuretic hormone (ADH), also called vasopressin,and (2) oxytocin.
If the pituitary stalk is cut above the pituitary gland but the entire hypothalamus is left intact, the posterior pituitary hormones continue to be secreted normally, after a transient decrease for a few days; they are then secreted by the cut ends of the fibers within the hypo-thalamus and not by the nerve endings in the poste-rior pituitary. The reason for this is that the hormones are initially synthesized in the cell bodies of the supraoptic and paraventricular nuclei and are then transported in combination with “carrier” proteins called neurophysinsdown to the nerve endings in the posterior pituitary gland, requiring several days to reach the gland.
ADH is formed primarily in the supraoptic nuclei, whereas oxytocin is formed primarily in the paraven-tricular nuclei. Each of these nuclei can synthesizeabout one sixth as much of the second hormone as of its primary hormone.
When nerve impulses are transmitted downward along the fibers from the supraoptic or paraventri-cular nuclei, the hormone is immediately released from the secretory granules in the nerve endings by the usual secretory mechanism of exocytosis and is absorbed into adjacent capillaries. Both the neuro-physin and the hormone are secreted together, but because they are only loosely bound to each other, the hormone separates almost immediately. The neuro-physin has no known function after leaving the nerve terminals.
Chemical Structures of ADH and Oxytocin
Both oxytocin and ADH (vasopressin) are polypep-tides, each containing nine amino acids. Their amino acid sequences are the following:
Vasopressin: Cys-Tyr-Phe-Gln-Asn-Cys-Pro-Arg-GlyNH2
Oxytocin: Cys-Tyr-Ile-Gln-Asn-Cys-Pro-Leu-GlyNH2 Note that these two hormones are almost identical except that in vasopressin, phenylalanine and arginine replace isoleucine and leucine of the oxytocin molecule. The similarity of the molecules explains their partial functional similarities.
Physiological Functions of ADH
The injection of extremely minute quantities of ADH—as small as 2 nanograms—can cause decreased excretion of water by the kidneys (antidiuresis). Briefly, in the absence of ADH, the collecting tubules and ducts become almost impermeable to water, which prevents significant reabsorption of water and there-fore allows extreme loss of water into the urine, also causing extreme dilution of the urine. Conversely, in the presence of ADH, the permeability of the collect-ing ducts and tubules to water increases greatly and allows most of the water to be reabsorbed as the tubular fluid passes through these ducts, thereby con-serving water in the body and producing very concen-trated urine.
The precise mechanism by which ADH acts on the collecting ducts to increase their permeability is only partially known. Without ADH, the luminal mem-branes of the tubular epithelial cells of the collecting ducts are almost impermeable to water. However, immediately inside the cell membrane are a large number of special vesicles that have highly water-permeable pores called aquaporins. When ADH acts on the cell, it first combines with membrane receptors that activate adenylyl cyclase and cause the formation of cAMP inside the tubular cell cytoplasm. This causes phosphorylation of elements in the special vesicles, which then causes the vesicles to insert into the apical cell membranes, thus providing many areas of high water permeability. All this occurs within 5 to 10 minutes. Then, in the absence of ADH, the entire process reverses in another 5 to 10 minutes. Thus, this process temporarily provides many new pores that allow free diffusion of water from the tubular fluid through the tubular epithelial cells and into the renal interstitial fluid. Water is then absorbed from the col-lecting tubules and ducts by osmosis, in relation to the urine-concentrating mechanism of the kidneys.
Regulation of ADH Production Osmotic Regulation. When a concentrated electrolytesolution is injected into the artery that supplies the hypothalamus, the ADH neurons in the supraoptic and paraventricular nuclei immediately transmit impulses into the posterior pituitary to release large quantities of ADH into the circulating blood, sometimes increas-ing the ADH secretion to as high as 20 times normal. Conversely, injection of a dilute solution into this artery causes cessation of the impulses and therefore almost total cessation of ADH secretion.Thus, the con-centration of ADH in the body fluids can change from small amounts to large amounts, or vice versa, in only a few minutes.
The precise way that the osmotic concentration of the extracellular fluid controls ADH secretion is not clear. Yet somewhere in or near the hypothalamus are modified neuron receptors called osmoreceptors. When the extracellular fluid becomes too concen-trated, fluid is pulled by osmosis out of the osmore-ceptor cell, decreasing its size and initiating appropriate nerve signals in the hypothalamus to cause additional ADH secretion. Conversely, when the extracellular fluid becomes too dilute, water moves by osmosis in the opposite direction, into the cell, and this decreases the signal for ADH secretion. Although some researchers place these osmoreceptors in the hypothalamus itself (possibly even in the supraoptic nuclei), others believe that they are located in the organum vasculosum, a highly vascular structure in theanteroventral wall of the third ventricle.
Regardless of the mechanism, concentrated body fluids stimulate the supraoptic nuclei, whereas dilute body fluids inhibit them. A feedback control system is available to control the total osmotic pressure of the body fluids.
Vasoconstrictor and Pressor Effects of ADH, and Increased ADH Secretion Caused by Low Blood Volume
Whereas minute concentrations of ADH cause increased water conservation by the kidneys, higher concentrations of ADH have a potent effect of con-stricting the arterioles throughout the body and there-fore increasing the arterial pressure. For this reason, ADH has another name, vasopressin.
One of the stimuli for causing intense ADH secre-tion is decreased blood volume. This occurs especially strongly when the blood volume decreases 15 to 25 per cent or more; the secretory rate then sometimes rises to as high as 50 times normal. The cause of this is the following.
The atria have stretch receptors that are excited by overfilling.When excited, they send signals to the brain to inhibit ADH secretion. Conversely, when the recep-tors are unexcited as a result of underfilling, the oppo-site occurs, with greatly increased ADH secretion. Decreased stretch of the baroreceptors of the carotid, aortic, and pulmonary regions also stimulates ADH secretion. For further details about this blood volume-pressure feedback mechanism.
Oxytocic Hormone
Oxytocin Causes Contraction of the Pregnant Uterus. Thehormone oxytocin, in accordance with its name, pow-erfully stimulates contraction of the pregnant uterus, especially toward the end of gestation. Therefore, many obstetricians believe that this hormone is at least partially responsible for causing birth of the baby. This is supported by the following facts: (1) In a hypophy-sectomized animal, the duration of labor is prolonged, indicating a possible effect of oxytocin during delivery. (2) The amount of oxytocin in the plasma increases during labor, especially during the last stage. (3) Stim-ulation of the cervix in a pregnant animal elicits nervous signals that pass to the hypothalamus and cause increased secretion of oxytocin.
Oxytocin Aids in Milk Ejection by the Breasts. Oxytocin alsoplays an especially important role in lactation—a role that is far better understood than its role in delivery. In lactation, oxytocin causes milk to be expressed from the alveoli into the ducts of the breast so that the baby can obtain it by suckling.
This mechanism works as follows:The suckling stim-ulus on the nipple of the breast causes signals to be transmitted through sensory nerves to the oxytocin neurons in the paraventricular and supraoptic nuclei in the hypothalamus, which causes release of oxytocin by the posterior pituitary gland. The oxytocin is then carried by the blood to the breasts, where it causes contraction of myoepithelial cells that lie outside of and form a latticework surrounding the alveoli of the mammary glands. In less than a minute after the begin-ning of suckling, milk begins to flow.
Related Topics