Chapter: Genetics and Molecular Biology: Biological Assembly, Ribosomes and Lambda Phage
Packaging the DNA and Formation of the cos Ends
Packaging the DNA and Formation of the cos Ends
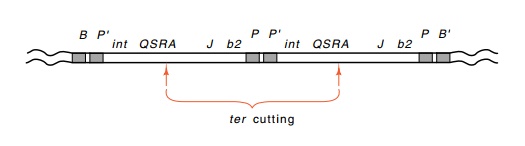
Up to this point, we have not considered the system
that converts circular lambda DNA, which is found
in cells, into the sticky-ended linear molecules that are found in phage
particles. The termini-produc-ing protein, the A gene product, generates the
required staggered nicks while packaging the DNA. The generation of these nicks
can easily be assayed by using a double lysogen in which both prophage are Int-
or Xis-. Normal phage excision is not possible from such a double
lysogen, but the ter system can clip
an intact lambda genome out of the middle of the two excision-defective lambda
genomes (Fig. 21.17).
In
experiments designed to study the effects on lambda of duplicating portions of
its DNA, a gratuitous duplication of the cohesive end occurred. A study of this
mutant has yielded appreciable insight into the packaging of phage DNA. The
starting phage for generation of the duplications was lambda deleted of a
sizable fraction of the b2 region of
the phage and containing an amber mutation in the red gene. These were grown, and selection was made for phage
containing duplications by selecting denser phage after separation according to
density on equilibrium centrifugation in CsCl. As expected, most of the
resulting dense phage contained duplications as shown by the following criteria:
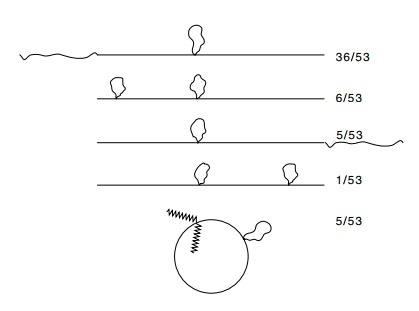
Figure
21.18 The structures of heteroduplexes
formed between a lambdacontaining a duplication on the left arm and a lambda
containing a duplication of the cos
site. The numbers on the right indicate the number of occurrences of the
various structures.
heteroduplexes
between strands from these phage and strands from wild-type phage contained a
bubble; the phage were denser than the parental-type; and, upon growth in Rec+
cells or in Su+ cells to suppress the red mutation, unequal crossing over around the duplication yielded
both triplications and phage lacking the duplication altogether.
If the
duplication phage were grown on cells unable to recombine and unable to
suppress the phage red mutation, no
change occurred in most of the duplication phage. One duplication phage,
however, was unstable under these conditions. At an appreciable frequency it
segre-gated phage lacking the duplication. Upon heteroduplex formation, an even
more startling result was found. The duplication could appear at either end of
the phage. To distinguish the ends in these experiments, the heteroduplexes
were formed between this strange phage and an-other phage containing a
duplication in the left arm. In all cases, one duplication bubble was formed in
the left half of the molecule and an additional bubble, that from the
duplication in the strange phage, was found near the right or left end.
A number
of types of heteroduplexes were found (Fig. 21.18). Two important conclusions
can be drawn from these experiments: the phage contains a duplicated cos site, but both cos sites need not be cleaved for the DNA to be packaged, and
packaging of the DNA proceeds from left

The
finding of a polarized left-to-right packaging of the lambda DNA is consistent
with the in vitro packaging
experiments investigating the nature of the DNA required for packaging. These packaging experiments found
that a molecule containing only a single genome’s equivalent of lambda DNA, a
monomer, was not capable of being packaged. Only polymers of the lambda genome
were capable of being packaged. Invivo,
the necessary polymers could derive from the rolling circle modeof DNA
replication or from recombination between circular molecules. The obvious
experiments were designed to discover the minimum DNA capable of being
packaged. The minimum is a lambda monomer con-taining a sticky left end and a
right cos end covalently joined to a
left cos end. These results all
strongly suggest that the left end of lambda ispackaged first and the right end
enters last. Since the tail is put on after packaging of the DNA, it is natural
to expect that the right end of lambda would be contained either within the
tail or just at the union of the tail to the head; indeed, when tailless lambda
phage that contain lambda DNA are isolated and are lightly treated with DNAse,
it is the right end of the DNA that is attacked. All of this strongly suggests
that upon infection, the right end of lambda should be injected first into the
cells.
Related Topics