Chapter: Ophthalmology: Ocular Motility and Strabismus
Ocular Motility and Strabismus: Basic Knowledge
Basic Knowledge
Ocular motility:
The movements of the eyeballs are produced by the follow-ing
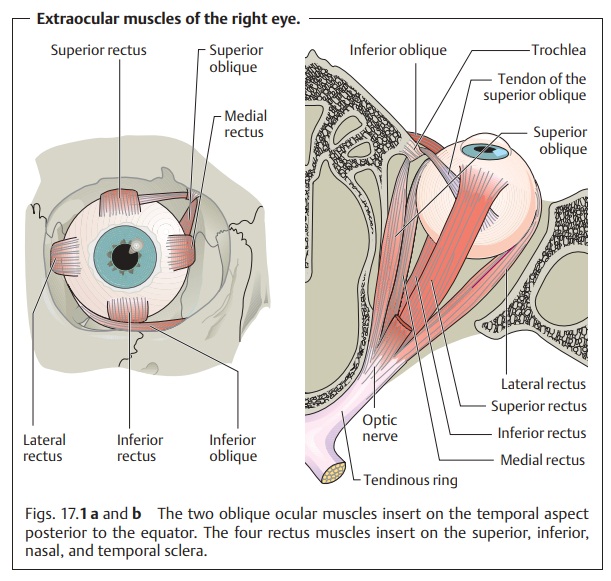
extraocular muscles (Fig. 17.1):
❖Thefour rectus muscles: the superior, inferior, medial, and lateral
rectus muscles.
❖ Thetwo oblique muscles: the superior and inferior oblique muscles.
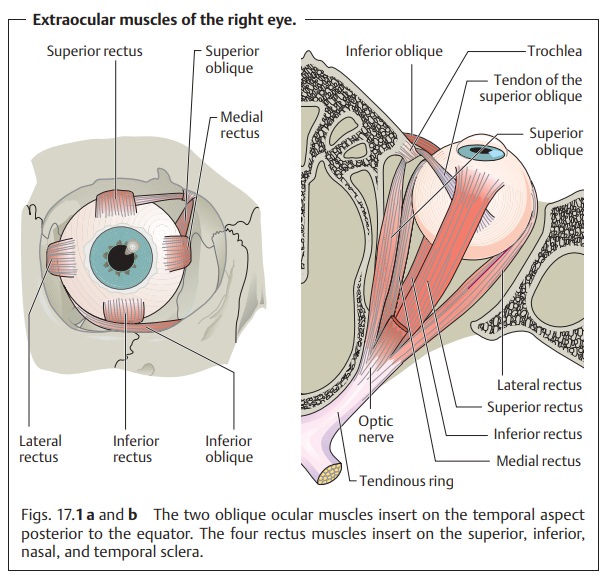
All of these muscles originate at the tendinous ring except for the inferior oblique muscle, which has its origin near the nasolacrimal canal. The rectusmuscles envelope the globe posteriorly, and their respective tendons insert into the superior, inferior, nasal, and temporal sclera. The oblique muscles insert into the temporal globe posterior to the equator. The insertion of the muscles determines the direction of their pull (see Table 17.1).
The connective tissue between the individual
ocular muscles is incor-porated into the fascial sheath of the
eyeball (Tenon’s capsule). Other impor-tant anatomic structures include the lateral and medial check ligaments comprising the lateral connections of the orbital connective
tissue and the ligament of Lockwood. This is comprised of the ligamentous structuresbetween the
inferior rectus and inferior oblique that spread out like a ham-mock to the
medial and lateral rectus muscles.
These anatomic structures and the uniform
nerve supply to the extraocu-lar muscles (like acting muscles have like nerve
supply) ensure ocular balance. Changes that disturb this balance, such as
ocular muscle paralysis that limits or destroys the affected muscle’s ability
to contract, cause stra-bismus. The angle of deviation is a sign of abnormal
imbalance.
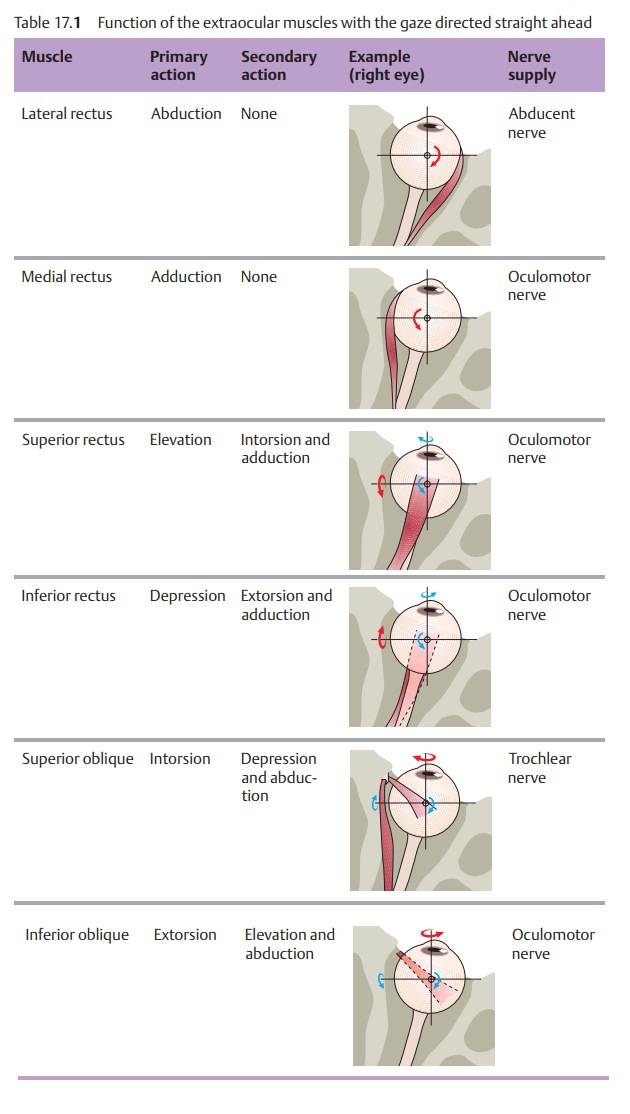
Direction of pull of the extraocular muscles: The
horizontal ocular muscles
The lateral rectus pulls the eye outward (abduction); the medial rectus pulls it
inward (adduction). All other extraocu-lar muscles have asecondary direction
of pullin addition to the primary one.Depending on the path of the muscle,
where it inserts on the globe, and the direction of gaze (Fig. 17.1), these muscles may elevate or
depress the eye, adduct or abduct it, or rotate it medially (intorsion) or
laterally (extorsion). The primary action of the superior rectus and superior
oblique is elevation; the primary
action of the inferior rectus and inferior oblique is depression. Table 17.1
shows the primary and secondary actions of the six extraocular muscles. A
knowledge of these actions is important to understanding para-lytic strabismus.
Nerve supply to the extraocular muscles:
The oculomotor nerve (thirdcranial nerve)
supplies all of the extraocular muscles except the superior oblique, which is
supplied by the trochlear or fourth cranial nerve, and the lateral rectus,
which is supplied by the abducent or sixth cranial nerve (see Table 17.1). The extraocular muscle nuclei are located in the brain stem on the
floor of the fourth ventricle and are interconnected via the medial
longi-tudinal fasciculus, a nerve fiber bundle connecting the extraocular
muscles, neck muscles, and vestibular nuclei for coordinated movements of the
head and globe (Fig. 17.2). Various visual areas in the brain control eye
and gaze movements. The location of the muscle nuclei and knowledge of the
visual areas are important primarily in gaze paralysis and paralytic strabismus
and of particular interest to the neurologist. For example, the type of gaze
paraly-sis will allow one to deduce the approximate location of the lesion in
the brain.
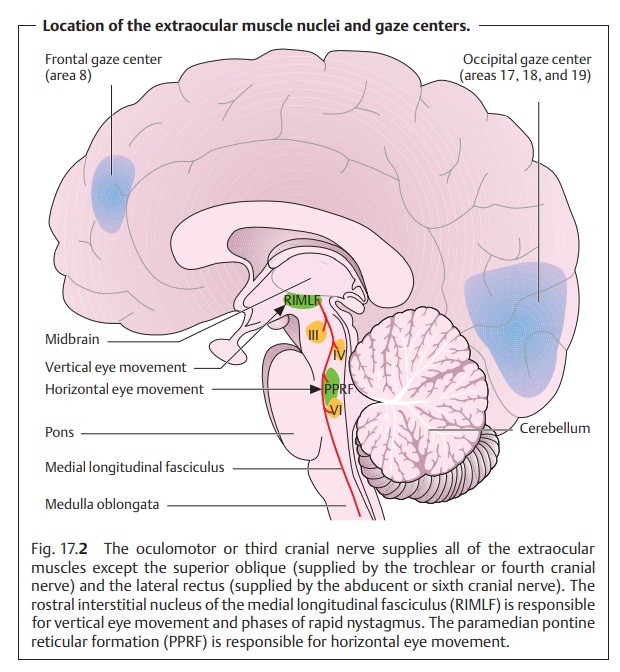
All extraocular muscles except for the
superior oblique and lateral rec-tus are supplied by the oculomotor nerve.
Physiology of binocular vision:
Strictly speaking, we “see” with our brain.The eyes are merely
the organs of sensory reception. Their images are stored by coding the stimuli
received by the retina. The optic nerve and visual path-way transmit this
information in coded form to the visual cortex.
The sensory system produces a retinal image and transmits this
image to the higher-order centers. The motor system aids in this process by directing both eyes
at the object so that the same image is produced on each retina. The brain can
then process this information into binocular visual impression. A person has no subjective awareness of this
interplay between sensory and motor systems.
There are three distinct levels of
quality of binocular vision:
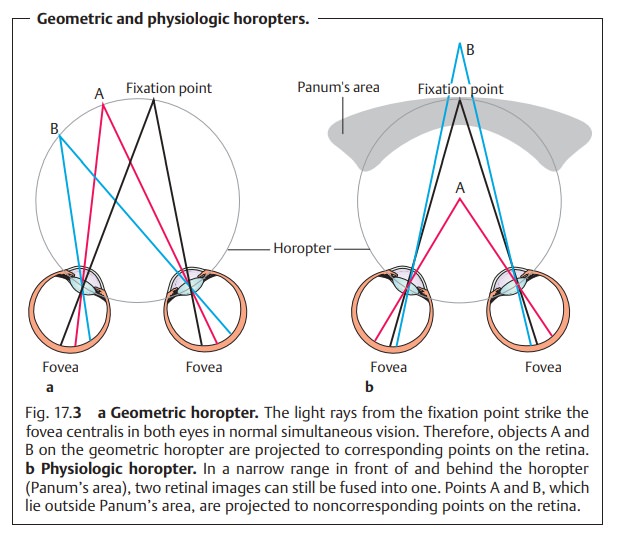
1. Simultaneous vision: The retinas of the two eyes perceive two images simultaneously. In normal binocular vision, both eyes have the same point of fixation, which lands on the fovea centralis in each eye. The image of an object always lands on identical areas of the retina, referred to as corre-sponding points on the retina.
Objects lying on an imaginary
circle knownas the geometric horopter
(Fig. 17.3a) are projected to these points on the retina. A different
horopter will apply for any given fixation distance. The images of both retinas
are therefore identical in normal binocular vision. This phenomenon may be
examined by presenting different images to each retina; normally both images
will be perceived. This is known as physiologic
diplopia.
Physiologic diplopia can be demonstrated by
placing two vertical pen-cils in a line along the subject’s visual axis, with
the second pencil approximately twice as far from the subject as the first.
When the sub-ject focuses on one pencil, the other will appear double.
2. Fusion: Only where both retinas convey the same visual impression, i.e.,transmit identical images to the brain, will the two retinal images blend into a single perception. Impaired fusion can result in double vision (horror fusionis or diplopia).
3. Stereoscopic vision (perception
of depth): This is the highest level of qualityof binocular vision and is
possible only where several conditions are met. For objects to be projected to
corresponding or identical points on the ret-ina, they must lie on the same geometric horopter. Objects lying
in front of or behind this circle will not be projected to corresponding points
but to noncorresponding or disparate
points on the retina. The result is that theseobjects are perceived as
double images (diplopia). However, objects within a narrow range in front of
and behind the horopter are fused into a singleimage.
This area is referred to as Panum’s area.
The brain processes noncor-responding retinal images within Panum’s area into a
single three-dimen-sional visual perception and does not interpret them as
double images (Fig. 17.3b). On the contrary, the brain uses the double images to
distin-guish differences in depth.
Related Topics